|
1 G' K) H4 i; Q" u+ D: U [新朋友] 点击上方天然气水合物" x7 O2 m. m/ `) Y2 g+ ~7 Z
9 [( z E9 U: d 关注 [老朋友] 点击右上角分享或转发本文本文导读在之前的文章中,我们对冷泉的基本概念以及冷泉碳酸盐岩. N6 d% L( K# `; ]( |- z$ B, d
" P5 d: T- J3 y5 ` 进行了简介(请戳下面的链接)。 % X. c" E9 R3 u
, j! b/ P: E( D2 f8 i8 h8 u3 e 冷泉——深海荒漠中的绿洲 * w8 e* S. W: q; {
今天的文章主要介绍冷泉生态系统。包括冷泉生态系统的化能合成作用、冷泉系统中的化能合成型生物群落以及冷泉生态系统对气候变化和地质活动的敏感性。
' @' g w3 k k% H0 p 
; ~' R) F+ H8 o- [8 C( v

( b6 b9 a- c, B# F8 n8 G" J
内容摘选自《“冰冻”的能源——联合国环境规划署对甲烷水合物的全球展望》 # I* |" F' t: F; R3 k; c2 m
海底冷泉释放出的甲烷或其他气体进入上覆水层。有些冷泉是与水合物相关的,而有些冷泉由于所处深度太浅而不能形成水合物。 6 X" S- f9 A0 r+ J, n
在这些冷泉中,甲烷和其他流体通过一些渗流运移到海底,这些渗流通道一般是通过过压、深层气藏渗漏、盐丘、泥火山口和地壳构造运动等形成的。
m3 W. ~" ?* y% V$ m) x$ g( F 甲烷冷泉可以用特殊的生物群落为标识,这些生物的新陈代谢以化能合成实现。因此,这些冷泉环境与洋中脊的热液排放是显著不同的。 / e- j5 |' j h- V3 k/ f
在海洋环境中,生物要利用从海底渗漏的甲烷中的“能量”有两种方式:缺氧氧化作用(AOM)和有氧氧化作用。
' }6 A3 m, t# z7 L; B* }/ C9 ] 缺氧氧化反应:细菌消耗甲烷的过程不需要氧气 4 L/ }& }% O5 U2 f2 O! [2 L+ e
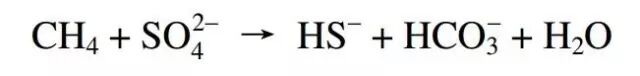
( r) s+ {+ \( L$ R. _
到达浅海海底沉积物中的甲烷有80%~90%会被微生物消耗掉。甲烷消耗的主要方式为缺氧氧化反应(图1 区域1)。缺氧氧化反应主要由两种微生物共同完成:食甲烷古细菌(简称ANME,即甲烷缺氧氧化细菌)和硫酸盐还原细菌。海水中含有非常丰富的硫酸盐,硫酸盐会渗入到沉积物中并且在甲烷的氧化过程中被消耗。图1中区域1的厚度与AOM反应的速率和甲烷的上浮通量有关,甲烷通量大时会薄一些(<10米),甲烷通量小时会厚一些。 ' I0 e- O/ R2 T0 L3 W7 `0 T; S
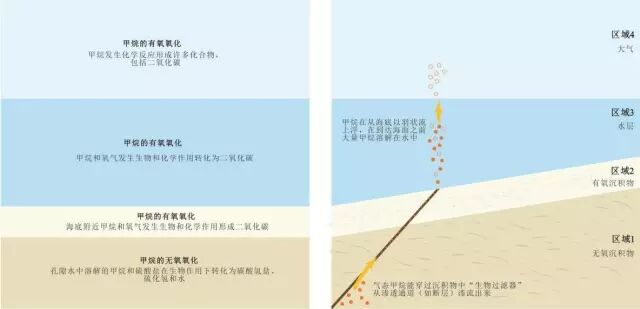
8 L) a/ ~' b6 W+ c3 A: t 图1 自然界中的甲烷消耗
# ?$ D: l2 L2 ~2 `5 j 注: 3 @3 D+ T! L0 L4 c3 F
近海底的甲烷水合物持续分解,释放出的甲烷溶解于地层水中。在甲烷穿过沉积物进入水层及大气层的过程中,大量地被化学反应和微生物作用消耗。
" U' U& I! r% N) ]3 ` [( J 如左侧图所列,作为延伸的化学合成食物链的一部分,地层水中溶解的甲烷会被微生物消耗。如右侧图所示,由于微生物作用只能消耗溶解在水中的甲烷,而气态甲烷能逃过沉积物中的微生物作用。甲烷气泡进入水层后能逐渐溶解于水,溶解后的甲烷能被有氧细菌消耗。这种生物甲烷过滤机制消耗掉了大多数的甲烷,剩余的甲烷气体进入大气。
8 D e. R+ ?4 B1 \2 v, p" a 上图没有按实际尺度绘制。一般来说,海洋中水合物存在的水深(区域3)为300~500米,甚至更深,区域2厚度为1厘米左右,区域1厚度为10米左右。
/ H9 S( X( U& s; H6 x" n; d! ] 一部分甲烷仍能逃过沉积层中的AOM反应。例如,在断层区域或泥火山处甲烷具有很大的通量时或者硫酸盐不能渗入沉积物时,甲烷会从沉积层的AOM消耗中逃脱。在这些区域,AOM不是一种绝对的海底甲烷过滤器,甲烷有时会穿过海底沉积物直接进入海水层。 3 a1 ]* e- c4 U' |' e3 m1 P, K7 @/ ]
有氧氧化反应:细菌消耗甲烷的过程需要氧气 ' p( A! f$ a' `, z; U- l4 K( G
甲烷的第二种消耗方式是有氧氧化反应。这个过程发生在同时含有氧气和甲烷的近海底沉积物中(见图1区域2),主要消耗AOM反应之后剩余的甲烷。甲烷的有氧氧化也是海水中甲烷消耗的主要形式(见图1区域3),尽管在一些地区应用了灵敏的放射性示踪技术,但是仍对伴生反应的相关过程缺乏了解。
1 J2 s* _9 u' Q7 }! _$ x# t; e) d 甲烷的有氧氧化是由食甲烷细菌进行的。甲烷是食甲烷菌唯一的能量来源,也是其机能碳的首要来源。被氧化的甲烷一部分转化为细菌生物质,剩余的都以溶解态的无机碳形式排放。AOM反应产生的主要无机碳是碳酸氢盐,而甲烷有氧氧化的主要产物是二氧化碳,二氧化碳会增加海水的酸性。
/ a; {; S$ D: i! T 在水体中,有氧氧化反应需要足够的时间和空间以供微生物有效地消耗甲烷。当甲烷扩散穿过水合物形成的区域(水深约300~500米)时,有氧氧化反应的效率很高。 1. 什么是化能合成?
. e& F& z$ F4 k1 @& k( B7 d 化能合成这个术语指一类特殊生物体的新陈代谢活动,这种新陈代谢利用氧化-还原反应中的化学能实现生物体生长。如果生物体生长所用的碳源为无机碳(二氧化碳),那么就称此类生物体为化能自养体。
& [( t5 W; t1 j7 R' [% H& z% ~ 化能自养生物与光合自养生物不同,光合自养生物以二氧化碳为原料利用光能进行光合作用实现生长,化能自养生物指的是细菌。动物(包括人类)以藻类、植物和其他动物为食,则称做异养型生物。 0 q+ _: C8 m- X9 V, B# j& K# d1 b' ]
化能共生动物的例子主要有大型管虫、蛤蜊、蚌等,它们在热液或甲烷冷泉系统等永久黑暗的环境里能够依靠化学能茂盛地生长。在深海中,这种生态系统能够完全不依赖阳光而正常生长。 $ V% S5 P7 i! u/ P8 Y& {' H
乍一看,这种说法好像是对的,因为作为这些生态系统的基础的主要生产者从无机物反应获取能量和碳,甚至整个过程不需要光。然而,高能分子如硫化氢的氧化反应需要氧气和硝酸盐。
J0 b; O( w s/ R/ f 这两种物质在远古海洋中很少见,只有通过光合作用才能产生氧气资源,而氧气进一步与还原性的氮气化合才形成硝酸盐。
7 l [5 ]9 V3 Q+ U8 V9 Y1 N 因此,现代复杂的化能合成生态系统,尤其是那些较高等的生物,不是完全与阳光没有关联的。它们会直接或间接地依赖光合作用产生的化合物。这种依赖性在冷泉明显加强。 0 L2 C7 M3 p8 N+ C$ C, A
冷泉系统中的甲烷或石油构成了化能合成食物链的基础。光合作用产生有机物质,有机物质又转化为化石燃料(甲烷或石油)。 " G2 L) o5 n; {, r

2 G6 K% E, w/ Y" m, m. u5 v 图2 墨西哥湾水合物丘
7 {/ W( V. V9 ?$ ^- _% ? 注:在活跃的甲烷冷泉之上、近海床位置能形成天然气水合物丘。上图为墨西哥湾水合物丘,水合物丘被少量石油染成橙色,部分覆盖有薄层沉积物(灰色物质)(Ian MacDonald提供)。 3 M& z8 T' q9 C. P% V; I
冷泉口形成的甲烷水合物会促使甲烷向更大范围的海底扩散,同时也会增加甲烷溶解于孔隙水中的量。溶解于水的甲烷更容易被化能合成型的生物群落消耗和利用,而气态甲烷通过非水合物稳定带的气体通道排放出来,从而逃过化能合成的生物群落。
# |- w' G+ ^" a; k' \6 q7 X 海底表面附近的天然气水合物可能还会促使沉积物中形成碳酸盐,因为碳酸盐是甲烷无氧氧化反应的产物。这些碳酸盐被侵蚀和暴露之后成为海洋生物的另一种生存环境。无论甲烷冷泉附近的海底表面是否有水合物,我们将不对甲烷冷泉生物群体间的差异进行区分,因为这些生物的适应性和生存方式几乎是相同的。
# b/ U3 r9 Z. ?3 M 发现与天然气水合物伴生存在的甲烷冷泉距今只有30年,因此对其研究仍处于初期。随着深海技术的进步,人们对这些冷泉系统尤其是大陆边缘冷泉系统的认识在逐渐增加。
- ^0 H- m7 q# U; P% k- X; k 尽管如此,我们知道这些生态系统在一些陆缘区域和构造活跃的海底区域普遍存在。对这些冷泉附近的化石(即暴露于甲烷冷泉海底的自生碳酸盐岩)的研究表明,这些海底生态系统已经存在了百万年之久。 2. 冷泉系统中的化能合成型生物群落
+ F; C) q. q1 S0 W: ~ 在甲烷冷泉处发现的化能合成型生物群落包括微生物、依赖硫化氢(AOM反应副产物)存在的动物以及直接消耗甲烷的动物。独立生存的化能合成型细菌包括硫细菌(如贝日阿托氏菌、辫硫菌属或硫珠菌等和有氧氧化甲烷型细菌。丝状硫菌有时能在沉积物表面形成大量白色或橙色的一层(图3)。 3 c% N4 _0 s- I( R

6 s5 `! Q! Z' T4 w) x2 \6 p, u
图3 与天然气水合物伴生的细菌簇 v# j0 i+ J6 S
注:太平洋东北部的Hydrate洋脊,白色和橙色的硫-氧化细菌簇覆盖在含有近表层水合物的沉积物上(Lisa Levin提供)。 2 {, [- |9 E" {
甲烷冷泉系统中的化能合成型动物有大有小,形式有丛状、堆积层状、礁状或单体生长,生长速率可能很快也可能异常缓慢。甲烷冷泉系统的动物群落可包含单细胞微生物(原生动物)或多细胞动物(后生动物)。 ! w, e$ l" R0 [ X4 j
其中大多数后生动物为无脊椎动物。许多都是以某种方式通过微生物活动与甲烷联系在一起而维持生存。
2 C/ v: W$ T5 `3 P ~ 常见的实例有海生管虫(图4A)、蟹(图4B、E)和各种蛤(图4C、F)。与非冷泉系统的深海动物群落相比,冷泉系统的动物群落相对较大。许多冷泉生物的消化系统都已部分退化或完全消失。 3 f0 U; K! F9 V
同时,它们为共生的化能自养型细菌提供居所,共生细菌则通过硫化物或甲烷的氧化反应为寄主回馈营养(图5)。 ' P4 Z' ?2 Y9 X, z& p
冷泉或冷泉系生物通过提供依附基质、分解化合物、夹带富有机质颗粒、供应微体原生生物或后生动物食物等方式,支撑着一个包含植食型、捕食型和以沉积物为营养的生物群落的繁荣景象。
6 k$ t1 o8 f$ H: `7 M9 x 此外,微生物群落在AOM反应中会沉积形成碳酸盐岩(石灰岩是一种碳酸盐岩),从而在冷泉附近形成结壳、岩石、巨砾甚至大型碳酸盐岩地貌。这些冷泉能维系高密度的蚌类、海底管虫和腹足软体动物。
+ Z; p4 o2 d* o6 i! v# G3 f* | 
N: f: @! s: A3 a( f. p 图4 冷泉生物实例 . u- N4 w5 G, }9 R
注: K! p% o# }$ M( g, t
甲烷冷泉生态系统的示例。A、C、E和F是化能自养动物,它们的能量来源于沉积物中微生物降解甲烷过程中产生的硫化氢。
! ^. d% ]$ u! `+ D1 ^+ j A:管蠕虫—巴勒姆氏瓣臂须腕虫; / D$ c Q5 b" @
B:在巨蛤和细菌簇区域中抱着管芯的海石蟹; % j5 V$ f" U" |/ S5 C& E( [
C:蛏螂—矩蛏螂; 5 m; r, c& ]+ ?! }
D:蜗牛—Neptunea amianta(一种绝迹的化石海蜗牛)和它们粘在岩石上的卵塔; + L7 _. ^; B$ U! b" d7 F$ i9 Q3 J
E:雪蟹—Kiwa puravida(雪蟹的一种,译者注),蟹螯上的“细毛”是丝状共生的细菌,雪蟹在富硫酸盐流体中挥动蟹螯培养细菌,再以细菌为食; 1 c" J$ X. J3 J; ` _
F:无齿蛤,在哥斯达黎加陆缘Quepos冷泉处(400米水深); " N0 [, W( {" D' ?
G:深海虾,位于哥斯达黎加陆缘第十二丘体(1000米水深) 0 ^# X# }' P* w: M# g1 v
(Greg Rouse和Lisa Levin提供)。
$ n, n7 p% A* |% F" M 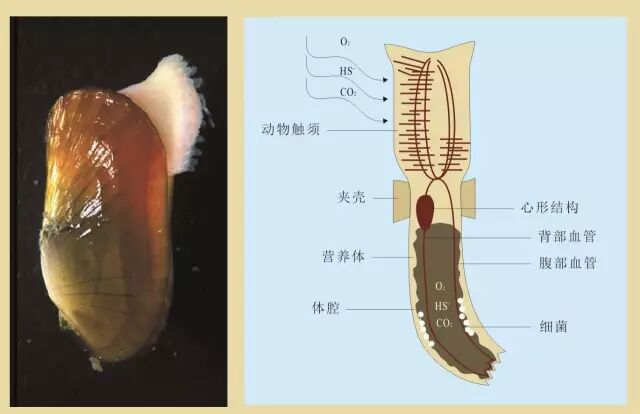
" N1 E. _8 T E* ^. k4 ~; x. _
图5 从AOM获取能量的共生生物组合
3 u: j( A1 O' a: z; m. g: \ 注:
! p* \% Z4 G' A, c9 I 某种管虫的形态结构(右侧)和蛤蜊与硫氧化细菌共生的图片(左侧,Greg Rouse提供)。管虫的共生体在其营养体内,营养体是一种特殊的器官。 4 g. C+ q, o. l \. F
管虫用动物须从周围水中吸收氧气(O2)、硫化物(HS-)和二氧化碳(CO2),通过内部血液流动将这些物质提供给共生体。蛤蜊将其共生体藏在它们的鳃里,从周围水中获取氧气和二氧化碳,并用脚从沉积物中获取硫化物。
6 V/ g' q4 s/ T+ n. u 因为这里所述的化能合成型的生物形态需要不同的甲烷和硫化物的化学平衡状态和浓度范围,所对应的流体化学状态和流速会形成具有一定特色的生物栖息区域。 3 q& y9 y+ [6 w8 K1 n
一般地,覆盖着硫氧化细菌席的沉积物与最强烈的液体和甲烷通量以及海底以下的天然气水合物有关;蚌和巨蛤层表示甲烷介于中等通量和高通量之间;蛤蜊和海生管虫表示甲烷通量较低且分散或者是天然气水合物埋藏较深(图6)。
^1 t- r4 j* g! M 这些联系已经在一些甲烷—冷泉环境得到证实。由构造型和基础型物种构成的微生物簇、层、丛和域等与沉积的碳酸盐岩一起形成了一种包含多种生物习性多样化的生物群落结构。 * U. Y. u- X4 `; A/ @& C4 q; [
这种结构对冷泉生态系统和大陆边缘的整体生物多样性都非常重要。冷泉系统中的生物很少直接与天然气水合物相联系。据文献报道,只有冰蠕虫Hesiocaeca methanicola(一种多毛虫)大类生物直接生活在甲烷水合物上。这种蠕虫具有相对较大的体型(2~4厘米),且群居密度很大(2500~3000个/平方米),发现于墨西哥湾的天然气水合物上。 0 q. {5 c8 E$ U7 a' p/ E3 Q: `
研究表明,冰蠕虫消耗与水合物相关的自生微生物,还发现蠕虫的活动会在水合物表面形成洼坑、制造小尺度水流,这些会促进微生物的生长并加速水合物的分解。冰蠕虫不仅生活在沉积物/水界面的天然气水合物上,还能生活在表层以下超过10厘米深处的天然气水合物上。
: z" T% i( k/ [2 W% w+ \5 R 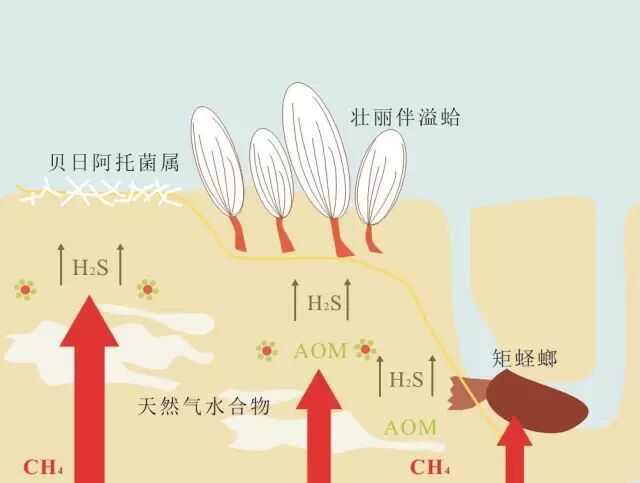
4 _' s) x2 G+ o" i' u
图6 化能合成系统 + C# U! F( b0 C' N% y) @% o
注: / d! z8 v5 S2 w2 A& g L
化能合成系统产生于不同流速的流体,包括甲烷的运移以及硫化物(甲烷a无氧氧化(AOM)过程产物)的运移,包含不同的动物群落。 3 z) }; C$ B" q4 ?( Y/ o
左侧:独立生存的硫氧化细菌簇(如贝日阿托菌属)生存于含有高通量流体的沉积物中。中间:巨蛤(如壮丽伴溢蛤)生存于含有中高通量流体的沉积物中。右侧:蛏螂(如矩蛏螂)生存于含有低通量流体的沉积物中。 . D0 Y5 U; C$ r7 F+ V
除墨西哥湾之外,尚未在其他区域发现与大块甲烷水合物样品有关的后生生物。尽管天然气水合物的存在提供了多种大量的附生细菌簇,而且这些细菌簇能导致附近底层内生物(生存在沉积物内的生物)的高密度聚居,但是在水合物脊区(Hydrate Ridge)裸露的甲烷水合物上并未发现后生生物直接聚居。 - E! M) u; R4 B7 ~2 v3 k0 X* h q
水合物脊区的细菌席之下的水合物实际上可能起到一层屏障的作用,可阻止某些蛤蜊、管虫和其他物种的进入。 3. 冷泉生态系统对气候变化和地质活动的敏感性
% [9 @! r1 L# K/ q 地质记录显示,气候变暖/变冷和海平面波动都会影响到甲烷水合物的稳定性、自生碳酸盐岩的形成、海底斜坡的稳定性以及冷泉生态系统的丰度。海底地震,如纽芬兰大浅滩地震,也能形成甲烷冷泉和化能合成型生物系统。 6 E/ {) u0 r7 n1 o, ~( H
目前,人们还不了解天然气水合物对持续气候变化的响应以及对化能合成生物群落的影响。水合物分解会增大甲烷冷泉的数量,从而创造出全新的生态系统,而水合物的快速分解和消失则可能会缩小现存冷泉生态系统的范围。
: Z; c( O$ F* ? 专业的天然气水合物(可燃冰)科普平台+资料中心 ( @' U4 t: R% M9 @5 s7 E
天然气水合物丨Gashydrate 1 R! h5 G9 X" j7 A8 S6 L
 ▲ ▲
; J( T0 C; B2 V: W5 o. o. P 关注天然气水合物及海洋地质领域近况与进展
$ v% e* R0 O0 o, W 特别致谢
9 R1 w& V$ _6 a8 ?' N8 G. X 感谢中国地质调查局地学文献中心为本文提供素材 2 ~4 m" i% z5 x! {
7 N/ ^3 F+ w0 \. F+ a
$ m z c l2 i) m* t& f1 j6 v5 e4 x& Y
; R! U9 e2 d2 P( L0 O' I) y @' A& c! d | 

