# ^1 K: P1 l* J% b 拓展探究问题海水的保护作用——海陆沉积环境差异对沉积物保存的影响往夕是何年?——如何确定钻探结果的年代海水古温度计——如何知晓过去的海洋温度说明:此为南师附中科学盒子中的拓展探究课题。 / e' s$ o! g" W4 c0 q4 \ `) z2 x- r
探究结果摘要
. z1 V# O+ x: {5 n4 a. X5 {# Y: A 在研究过去海洋生物地理分布、全球气候变化时,沉积物能提供十分有用的线索。由于海洋的特殊环境,海洋沉积物在洋底经过长时间积累而成,包含了近于连续的,横跨百万年的时间序列,为我们提供了大量关于过去地球状况的信息。
5 z) X! ]+ s; M5 l6 p0 V9 a. j 地质年代包含相对地质年代和绝对地质年代两重含义。相对地质年代可以通过地层层序或生物层序间接分析得出,绝对地质年代可以通过同位素法测定。地质年代表便是结合了这两者建立起来的。
5 l) E, A" L e; n' S' S) w$ F, F 过去某一片地区的海洋温度可以通过微体古生物法或稳定同位素法定性或者定量地得出。得知海洋温度之后可以推知过去的洋流等海洋信息。 . p0 P4 e' R+ f3 |5 v
. C% _! A J0 S3 r* R* ~! `3 s6 }
海洋沉积物中深海沉积物所蕴含的信息最为丰富。 2 u2 Q. ]3 T& ^
浊流是一种携带大量碎屑物质而十分浑浊的水体,其大多形成于大河河口外或大陆坡上坡度较大且有大量碎屑物的地段。当海底有地震或火山喷发等外因触发时,海水携带大量碎屑物质,在重力的作用下沿海底斜坡快速而下,直至深海地形较为平坦的地方堆积下来。
8 }/ j! x% D/ O! k8 j 深海是水深超过2000m的广大海域,面积占地球表面积的59.5%以上,是大洋的主体部分。深海的水温和盐度都不受大陆影响,具有独立的洋流系统。
5 X7 `$ A1 m- X) w" y 由于远离大陆,深海沉积物来源远不及浅海,以颗粒细小的泥质和化学、生物物质为主。浊流携带者小颗粒的泥沙和植物碎屑、浅水生物遗体以及其他浅水沉积物一起来到深海,与来自于深海的沉积物一同构成生物软泥和红色黏土(含火山碎屑较多的黏土沉积物,在强氧化作用下显红色)。生物软泥在较为稳定的环境下像这样经过百万年的时间跨度沉积下来,故海洋相较于不断变化的陆地来说更加有利于沉积物的保存,并且更有价值。 6 e. c6 x6 f- [/ V* N$ C3 v
海水沉积物为我们提供了大量关于过去地球状况的信息。通过采集设备采集岩芯(圆柱状沉积物和岩石),并认真分析和解释,地球科学家就能推断过去的环境条件,比如海表温度、营养供给、海洋生物丰富性、大气运动状况、洋流形式、火山喷发、重要灭绝事件、地球气候变化、构造板块运动状态。事实上,目前我们所得到的关于过去地球地质概况、气候、生物方面的认识,都是通过对古海洋沉积物的研究得出的。 " ]+ P( u" G% f" r$ E% ?% f
. j, ?" ?1 w: a6 ]* Z: d3 U 地质年代指地质体形成或地质事件发生的时代。它包含两层含义:地质体形成或地质事件发生的先后顺序,称为相对地质年代;地质体形成或地质事件的发生距今有多少年,称为绝对地质年代。 + ^ L4 m7 y& z
1. 相对地质年代的测定方法0 u& Q6 X" G0 u* J; t
1.1 地层层序律 7 |8 G( P& ~# z8 q
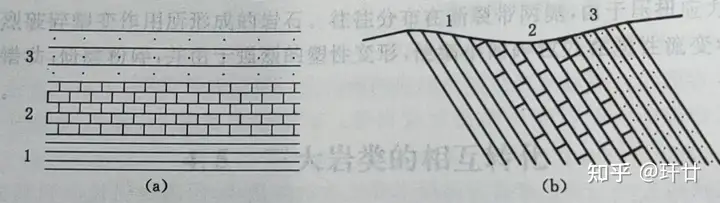
地层形成时的原始产状是近于水平的,老的地层在下,新的地层在上,这就是地层层序律。如果地层因构造运动而发生倾斜,则倾斜面上的地层新,倾斜面下方的地层老(如图1)。
5 N$ ]/ }% T6 H4 q- U/ D 图1 地层的相对新老关系 (a)地层水平 (b)地层倾斜地层层序律的最大不足是,地层原始产状经常被后期的构造运动改变,发生倒转,上下颠倒。 1 @6 {; n, v; m: I: ?5 O
1.2 生物层序律
; v+ S8 {8 R" K3 L, J 地层层序律只能确定同一地区相互重叠在一起的地层的新老关系,要对比不同地区的地层之间的新老关系,常常利用保存在地层中的古生物化石来确定。大体上,年代越老的地层中的化石更加原始,形态结构更为简单;年代越新的地层中化石更加高级,形态结构更为复杂。不同时代的地层中所含的生物化石组合不同,相同时代且原先海洋和陆地相通的地层中有相同或相似的古生物化石。这就是生物层序律。
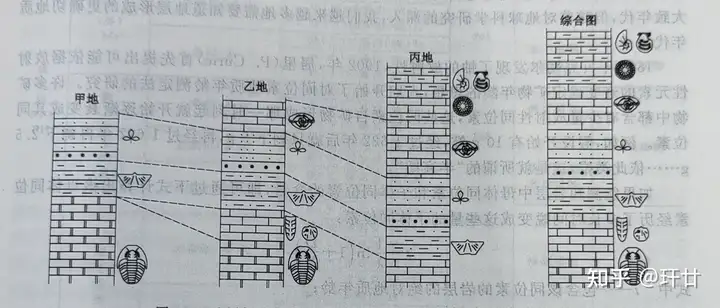
& y* O- B7 F7 R: G5 B: w 根据生物层序律不仅可以测定地层的现后顺序,而且还可以确定地层形成的大致年代。下图展示了如何利用生物层序律对比甲乙丙三地地层的新老关系,并通过对比还原该地区完整的地层层序,建立起统一的综合地层柱状(如图2)。
7 u0 E$ }6 u; g- j* X1 v& t0 i 图2 地层划分与对比及综合地层柱状图值得注意的是,有些生存时间较长的生物不适合用来确定地层年代(例如舌型贝,从寒武纪生存到今天,形态特征没有明显变化)。生物层序律的最大不足是,火成岩、深变质岩中没有生物化石;有了生命以后的沉积地层和软微变质的变质岩中才有生物化石。 * D( b# e) [( d1 {; y4 m4 p
1.3 切割律 # S+ ]. U7 D) m) ?6 ]" i
上述两条准则只适用于确定沉积物或层状演示的新老关系,对于块状产出的火成岩或变质岩则不适用。这时,他们之间的新老关系可以依据切割律来判定(如图3),即入侵者年代新,被入侵者年代老;切割者年代新,被切割者年代老;包裹者年代新,被包裹者年代老。
. s! d3 i9 M7 E5 [3 G 图3 运用切割律确定各种岩石形成顺序示意图2. 绝对地质年代的测定方法:同位素法$ t/ g0 D9 A$ g0 l( [" S
相对地质年代只能确定地层的新老关系,即使运用生物层序律也只能了解地层形成的大致年代,而同位素法却能够得知更确切的地质年代。许多矿物中都含有少量放射性同位素,这些母体同位素自矿物形成的那一刻起就开始逐渐衰变成其子体同位素。只需要知道有多少母体同位素衰变为了子体同位素,便可以反推矿物形成的时间。
4 a/ b8 ^1 Q" z$ J2 C- t, U 同位素法的原理是基于放射性元素都具有固定的衰变常数( λ\lambda )(即每年单位质量的母体同位素能产生的子体同位素的质量),且矿物所含放射性同位素蜕变后剩下的母体同位素含量( NN )与蜕变而成的子体同位素含量( DD )均可测出,即根据公式 :
- v9 x! g6 {0 ^7 } t=1λln(1+DN)t = \frac{1}{\lambda}\ln(1+\frac{D}{N}) \\
( y b N( [' H5 o1 q* u 就可计算出该放射性同位素的年龄( tt )。它就是包含该放射性元素的矿物的形成年龄,称为矿物的同位素年龄,即相当于包含该矿物并和该矿物同时形成的岩石具有的年龄。
- R. v. E. Y( p' [ 同位素测定法也存在一些不足,例如有些沉积岩不含有与沉积作用同时形成的放射性同位素,而且放射性同位素在一般矿物中含量很低,对精度要求很高,故测定难度大,容易造成人为误差。 / T; s( m! J3 {/ F' l
3. 地质年代表
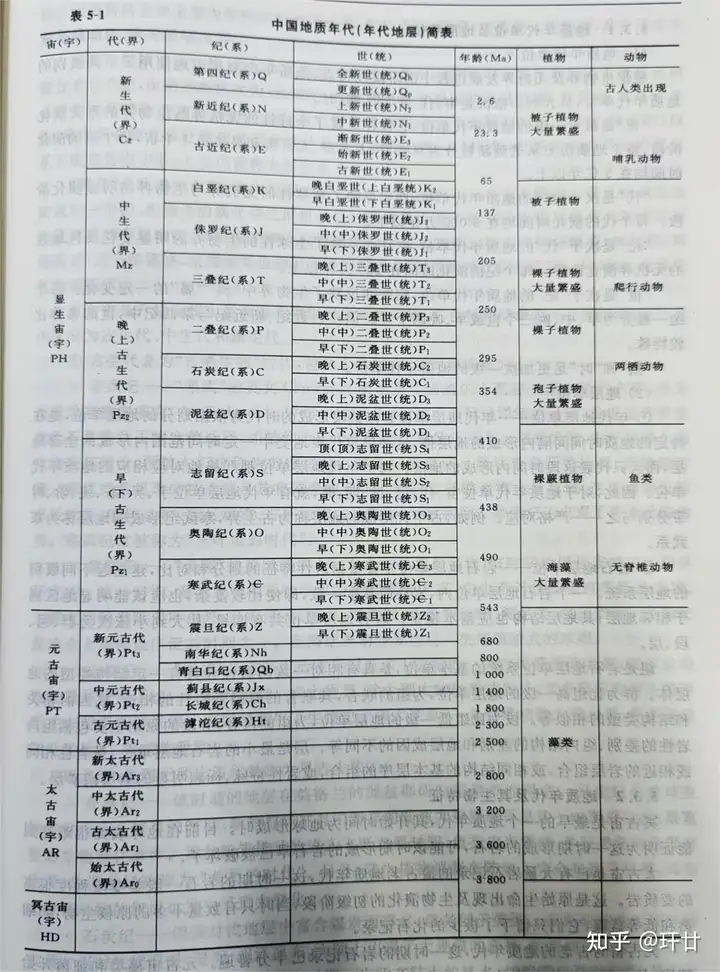
; S' U- K# \, H- Y7 z/ e4 } 地质学家通过对全球各地的地层进行对比研究,发现地层的生物组合特征、岩性特征等均呈阶段性,且大阶段里面包含小阶段,这种阶段性是全球统一的。在此基础上,结合同位素地质年龄测定法,对漫长的地质历史进行系统性的编年与划分,便制定出了一个在全球范围内普遍参照对比的年代表,即地质年代表。(如图4) ! N' E0 n' W; w4 Y4 _6 d( R
图4 地质年代表- u5 A/ y2 W& `- {* d4 ?
过去某一片地区的海洋温度可以通过微体古生物法或稳定同位素法定性或定量研究。
o. O9 q7 T1 @+ s6 g" \4 q 1. 微体古生物法$ A, O. j) Z6 H# @# C1 m1 U% ]& B
对沉积环境的变迁,生物的反应最为灵敏。能够作为水文标志的生物须有不能快速移动、数量丰富等特征。浮游有孔虫最能符合上述条件,事实上它已经成为古海洋学研究中指示表层与近表层水体特征的主要标志,被誉为“海流的镜子”;钙质超微化石在这方面也能起相似的作用,有时还可利用翼足类化石。为了解底层水的水文特征,通常使用底栖有孔虫,有时也用介形虫化石。此外,在钙质为题花式缺乏的情况下,硅质微体化石便成为古海洋学研究的主要材料,例如放射虫与硅藻。
3 \0 U8 U: L6 ~: z0 @ 1.1 标志性种和标志性组合
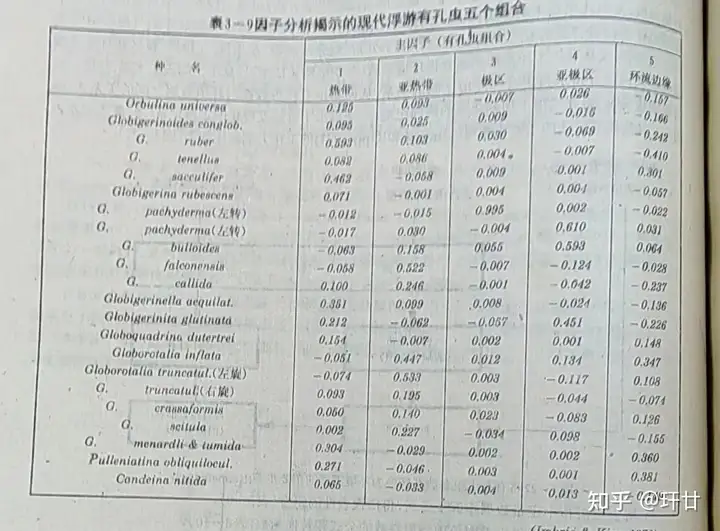
* c1 D5 a0 v+ B) y1 J0 y 大洋表层与深部的水团(指物理化学性质稳定,变动状态保持一致的一部分水体),由于各自的形成机制、源地深度、温度、盐度等具体情况不同,可以形成特性各异的物理、化学环境,构成了不同的生态条件,形成各自的“生态水团”。它们分别控制着一定的生物群落,具有很强的专属性。这种隶属关系很明确,故可以通过分析现代各种水团的标志性种与标志性组合,再根据钻探得到的岩芯中的化石,反推出岩芯中沉积物形成时的古水团的种种特征。(如图5)
3 k w$ Z. j" x3 Z+ } 图5 现代浮游有孔虫主要种的温度分布1.2 非种方法
( Z" H m$ C+ m+ _" y' V) [ 标志种和标志性生物组合固然是微体化石用于古海洋学的基本方法,但是由现代属种组成的化石群,只有在晚近地质时期才能遇到,因而在较老的地层中直接与现生种比较的方法受到一定限制,以化石群的群落结构或化石种群的形态特征为基础的非种方法,不受地质时代与属种演化限制,具有更加广泛的应用范围。 1 {6 t* n9 @+ |( F ]
1.2.1 群落结构 / V4 r; i# H( @% o3 `! j
化石群有许多指示环境的特征,其中以分异度最为重要,即生物群落中分类单元(如属、种)多元化的程度。其计算方法中最简便的便是统计一标准化样品内化石群中分类单元的数量。 - ]$ j# W# F/ N7 C( d/ o
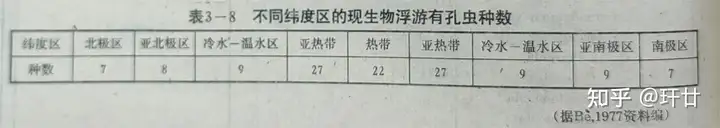
分异度之用于古海洋学,其根据是现代生物群的分异度与纬度、温度、盐度、深度等环境因素有密切联系。在开放性大洋中,盐度变动微弱,但纬度、温度对分异性影响甚大。以有孔虫为例,现代有孔虫共计有30~50种,在不同纬度出现的种数向两级递减(如图6)。 ' v. `: I3 u5 L% V2 V9 X6 V O3 t
图6 不同纬度区的现生物浮游有孔虫种数故通过分析现代各种生物分异度和温度的关系,再根据沉积物样本中生物群落的分异度,可以反推出沉积物形成时的环境特征,如温度等。
2 ~1 I* [+ i" T# ` 1.2.2 形态特征 8 R8 U. y9 Y; u5 k8 o
大洋中,微体生物的生态表型也往往受到环境条件的制约,具有一定的指温、指纬意义。这包括壳体的形态、壳径、壳面空隙率、壳体转向、壳口大小等形态结构的变量。故通过研究现代微体生物的生态表型与温度、纬度的联系,再根据沉积物样本,可以反推出沉积物形成时的环境特征。 ) }$ z9 D3 |, }% ]5 e( ?; I9 U
1.3 定量方法 , b" `6 m2 F1 @* N8 [. R" e
上述种种古生物学方法,实际上都只能提供古海洋学的定性结果,如揭示水团的属性、水温的相对高低,但并不能定量地得到温度等具体数值。为进行古水温等参数的定量研究需要求助于其他方法。目前采用的主要有两种:一是用生物壳体求氧与碳同位素比值的方法(后文中会讲到),这属于地球化学的范畴;另一是用转换函数法处理微体化石群的定量分析数据,即用数理统计的技术来确立生物组合与古温度之间的定量关系。 , h2 J& ]2 n. j1 |7 j; a
所谓转换函数是指两类数据之间的关系式,这里指微体化石群的数据与当时环境数据之间的关系式:
+ h% w0 d% A3 c; Q" Y Ta=f(P1,P2,⋯,Pn)T_a=f(P_1,P_2,\cdots,P_n) \\ 3 K! d2 C4 B2 _/ B
式中 TaT_a 是温度值, P1,P2,⋯,PnP_1,P_2,\cdots,P_n 分别代表微体化石群中各个种的相对丰度(百分值)。 + _" D j6 S# e
此式的含义是通过各个种相对丰度间复杂函数的计算,求出所需的古温度值。 3 h* N$ u8 J- j0 r- K0 T) x
1.3.1 最简单的方法
& L- [# Q. b; U4 M# t 可以想到一种最简单的关系式,例如采用各个种最宜温度的加权平均数的方法。当需要求出某个样品所反映的古温度时,首先求出每个种的百分含量,然而用加权平均数将生物丰度的信息转换为对应的古温度值: % T- P! }- p/ S8 k: t/ K
Test=∑PiTi∑PiTest = \frac{\sum P_iT_i}{\sum P_i} \\
. k$ Y! n- h& r0 ?! U: l 式中 TestTest 为所求的温度估算值, PiP_i 为第 ii 种的百分含量,TiT_i 为第 ii 种生存的最宜温度。
: X- E+ q6 t! j9 H" g% C2 K! T0 F8 O 数十年前,Berger提出用浮游有孔虫定量测算古温度的,正是这种方法。如果想要测得精确些,可以考虑一个种的温度分布范围,而不只是其最宜温度值,但过程相应地会复杂些。 V4 \; x6 n& F; T2 q3 A0 A" H |3 ?
1.3.2 古生态学转换函数
5 E, d# ^5 y- }& q" ^3 X' n 在了解这个方法之前,需要先了解两种数据分析手段:因子分析和回归分析。 & j3 u- _: `. q8 ]6 E& f( y# ~, x
因子分析是从变量群中提取共性因子的统计技术。(具体介绍请看这篇文章) " f5 f E/ ~0 {
回归分析是指的是确定两种或两种以上变量间相互依赖的定量关系的一种统计分析方法。以接下来要用到的多元多重线性回归分析为例,它需要找出多个因变量和多个自变量的关系,最终得出了三个线性的关系式:
9 z/ p! E0 j7 b' l' o3 n Ts=19.7A+11.6B+2.7C+0.3D+7.6Tw=23.6A+10.4B+2.7C+2.7D+2.0S=2.0A+1.9B+0.8C−1.6D+33.8 Ts=19.7A+11.6B+2.7C+0.3D+7.6 \\ Tw=23.6A+10.4B+2.7C+2.7D+2.0 \\ S=2.0A+1.9B+0.8C-1.6D+33.8
$ |1 S% z1 O# C4 B1 {8 O7 V: B 古生态学转换函数由美国的Imbrie与Kipp发明,是迄今为止最好的古温度定量方法之一。它采用因子分析和回归分析技术从化石群的定性分析中提取温度的信息,其主要步骤以这两位科学家的工作为例简述如下: 将表层沉积中定量数据作因子分析,划分出热带、亚热带、亚极区、极区和环流边缘等五个组合(如图7)从现代大洋参数图集查得环境参数(冬季平均表层水温.支季平均表层水温、盐度),再通过多次回归分析得出求取各项环境参数的公式和转换函数(如图8) 将钻孔剖面中逐层的化石数据按上述有孔虫组合进行分解,即: udc=FdcVu_{dc}=F_{dc}V
% T& O U7 b3 |5 g1 X- q6 v 式中 FdcF_{dc} 代表钻孔中逐层的有孔虫定量数据, VV 为有孔虫组合的描述矩阵(即图7), udcu_{dc} 为所求化石组合的数
4 G8 w& H# N3 w) O7 ] ~ 4. 用转换函数(即图8)对钻孔中化石组合值( udcu_{dc} )作古环境推算( EdcE_{dc} ,包括夏、冬季古温度和盐度) ! Z& W, `' _- R8 r7 g
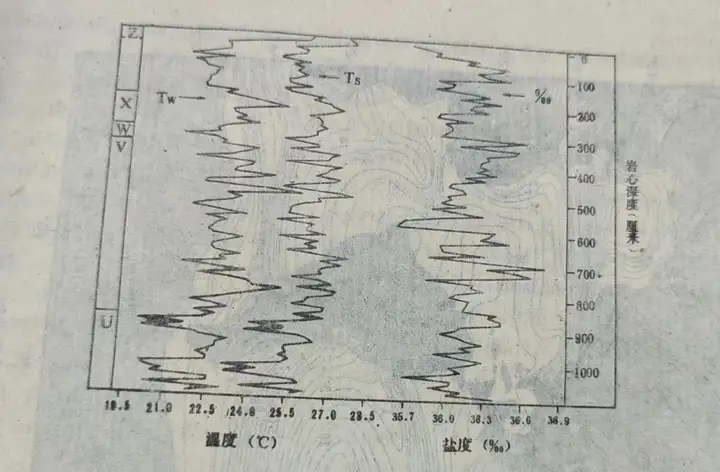
所得结果如图9。 , m2 ?: S4 g% f
图7 因子分析解释的现代浮游有孔虫的五个组合图8 用浮游有孔虫组合推算环境的公式图9 柱状样品中浮游有孔虫反映的古温度、古盐度变化2. 同位素法
5 [. [% R/ g! D! n: ^) k 在自然界中,氧元素由 16O^{16}O (占99.76%), 17O^{17}O (占0.04%), 18O^{18}O (占0.20%)三种稳定同位素组成,碳元素由 12C^{12}C (占98.89%)和 13C^{13}C (占1.11%)两种稳定同位素构成。为了说明某元素同位素组成的特征,通常用两种同位素的含量比值来表述,如 18O/16O^{18}O/^{16}O ( 17O^{17}O 含量极微忽略不计), 13C/12C^{13}C/^{12}C 。
4 w, ~+ b( {& K+ n' _: @9 m 从元素的原子结构来看,由于同一元素诸同位素的质子数及核外结构相同,因此它们的化学性质极为相似,但它们的原了量又各不相同,因而在某些物理一化学过程中(如蒸发、沉淀、凝聚、结品、熔化、扩散等)却能显示出微细的差别,谓之同位索效应。这样,当物质间发生相互作用或转化(如碳酸钙与二氧化硅从水中汇淀,大气与海水交换)时,就会导致同位素间的交换与再分配,轻、重同位素在不同物质中将相对富集,同位素组份比值也随之发生微小的变化。这种在物理-化学过程中造成某同位素在两种物质中分配上的差异,称之为同位素的分馏作用。但是,在自然界中,这种变化是极其微弱的,难以直接测出其绝对比值,需将变化后的比值与某种人为确定的标准样品的固有比值加以比较,再将两者偏差的千分率作为同位素组份的的量值 δ\delta ,其代表式为:
% B) f3 p( L+ j 测定样品的比值标准样品的比值标准样品的比值测定标准标准测定标准标准δ=测定样品的比值−标准样品的比值标准样品的比值×1000δ18O=(18O/16O)测定−(18O/16O)标准(18O/16O)标准×1000δ13C=(13C/12C)测定−(13C/12C)标准(13C/12C)标准×1000\delta= \frac{测定样品的比值-标准样品的比值}{标准样品的比值}\times1000 \\ \delta^{18}O = \frac{(^{18}O/^{16}O)测定-(^{18}O/^{16}O)标准}{(^{18}O/^{16}O)标准}\times1000 \\ \delta^{13}C = \frac{(^{13}C/^{12}C)测定-(^{13}C/^{12}C)标准}{(^{13}C/^{12}C)标准}\times1000 \\ : n* k- ]8 @! t; R5 E5 Q3 _$ B
2.1 氧同位素
! X! l) X7 J. [+ t( P* C 早在1947年,科学家Urey就指出了二氧化碳-水-碳酸盐系统中氧同位索的分馏作用与海水温度间存在着相关关系,成为利用氧同位素测定古水温的出发点。当碳酸钙从水溶液中沉淀出来(进入生物壳体)时,相互间发生同位素交换反应,反应方程式为: ' L3 f! U0 Z- g& i; C
1/3{CaCO_3}^{16}+{H_2O}^{18}\leftrightharpoons 1/3{CaCO_3}^{18}+{H_2O}^{16} \\
5 V: U# {" |4 } 当反应达到平衡时,其平衡常数与温度间有确定的关系,即碳酸钙的氧同位素组成是温度的函数。当温度升高时,相对较轻的 ^{16}O 由于有较高的活性,易于迁移,在同位素交换反应中将优先被吸收进生物壳体中,致使 ^{18}O 含量相对减少,故 \delta^{18}O 值随温度的上升而下降。 & P* v+ j; | q) f' f
研究表明,同一地区的浮游有孔虫的 \delta^{18}O 值随季节和海水深度变化,充分证明了水温的控制作用。所以通过研究现代样本的\delta^{18}O值,并与过去样本的\delta^{18}O值进行比对,可以推知样本形成时的温度。
4 x8 ?. e9 E0 g( C* {; Y0 q( b 值得注意的是,因为 \delta 是可以精确算出的数值,所以通过回归分析等手段可以得出 \delta 与 T (温度)的关系式,进而算出较为精确的温度值。氧同位素的分析研究可以帮助查明地质时期海水温度变化趋势、揭示全球气候变化规律、建立大洋水柱垂向温度梯度剖面等。不过\delta^{18}O值容易受到如冰期效应、盐度效应等的影响,研究时需要根据实验排除干扰。
8 [/ g9 t4 J2 Y7 X 2.2 碳同位素 # ?' K, [2 Q4 l6 O; h1 b6 L$ t
海水中 \delta^{13}C 值的变化主要与生物的生命活动(生长、繁殖、光合作用、新陈代谢)密切相关,其中尤以光合作用最为重要。在光合作用下,植物优先吸收较轻的 ^{12}C ,使植物体内的\delta^{13}C值变小,而海水或者大气中\delta^{13}C值相应升高。这种光合作用的影响是十分迅速且有效的:一昼夜内,海洋中\delta^{13}C值的变化可达±0.1‰,陆地上甚至可达2‰。故通过与氧同位素类似的方法,可以推知过去某一地区的较为准确的海洋温度。 , {3 f0 `) p0 U2 e/ s J1 C
由于影响碳同位素成分变化的环境因素复杂多样,碳同位素的分析和解释比氧同位素要复杂很多,但也正因为如此,一旦将它们逐一研究清楚之后,那么它不仅可以用来推断海水古温度、古盐度、冰期效应,还可以用作再造古生产率、大气成分、水体性质以及日照强度等的依据。这使得碳同位素分析研究的重要性在氧同位素之上。
8 Y# m/ Z9 L6 a0 g$ Z 参考^舒良树.普通地质学[M].地质出版社,2010.^Alan.P.Trujillo.海洋学导论[M].电子工业出版社,2017.^同济大学海洋地质系.古海洋学概论[M].同济大学出版社,1989.
- q! q: e V" [9 o. L8 r5 S' c! z
: T( @; ~; |% @+ r; i/ C
) L( R- P j3 R' ~2 P5 z2 e0 Z/ B
3 \! |4 h' t F; J, d I/ @
|