|
. L2 V; U3 ^6 Y) ?3 G+ p, b 
 6 n: l$ ~- S) |0 j1 j# v
6 n: l$ ~- S) |0 j1 j# v 【论文题目】环境DNA揭示海洋生物群落季节性变化与潜在相互作用(“Environmental DNA revealsseasonal shifts and potential interactions in a marine community”)
% T `2 ]: G5 h9 U5 \, q 【期刊名称】Nature Communications,(2020)11:254 * n y9 j( C0 ?% H
【第一作者】Anni Djurhuus
% |& t. O+ R* I& m7 B2 s3 t, Q4 D 【通讯作者】Alexandria B. Boehm & Mya Breitbart
2 o# e. U3 M W0 e2 K( _# |& B 【全文链接】https://doi.org/10.1038/s41467-019-14105-1 3 q, d5 O* {. e
亮点 + q* R, y+ S6 u: w1 |3 ?9 D( |
1、评估了海洋中不同生物类群相对丰度比例的变化,并分析微生物到哺乳动物等生物之间的关联;
/ j9 y' P k3 i 2、阐明了群落动态和环境因素之间的关系,即不同生物类群的丰富度和多种环境变量(包括海面温度、叶绿素a等环境和生物因素)之间的关联;
; F" n& z8 l+ L% {% ^. T) }* b 3、发现了跨越不同营养级的共生生物群与环境参数的变化直接相关,明晰了群落对环境的潜在响应,并强调了共生生物和营养级间的相互关联。
, L4 V' t' u: z, ?3 A# J/ Z/ [ 摘要 . q$ B; A. y. }" |
环境DNA (eDNA)分析可以同时检测不同营养级的生物类群,从而提供与生态系统变化相关的复杂生物相互作用的关键信息。本研究利用eDNA的多位点扩增序列对加州蒙特雷湾国家海洋保护区的海水样本进行了为期18个月(2015-2016年)的生物多样性调查。由此产生的数据集包括663个分类群(在科或更高的分类级别),范围从微生物到哺乳动物。我们推断了群落组成的变化,揭示了类群间的相互作用,并确定了这些群落与环境属性之间的相关性。群落网络分析提供了预期的捕食者-被捕食者关系、营养联系和季节变化的证据,跨越生命的所有领域。我们得出结论,基于eDNA的分析可以提供有关海洋生态系统动态的详细信息,并识别敏感性的生物指标,从而为制定生态系统的保护策略提供重要信息。  ; F" n2 `2 F0 B; [ ; F" n2 `2 F0 B; [
1. 引言
J* s/ c3 ]$ D) P1 X8 l 由于无法同时评估不同生物在多个营养级的丰度和分布,所以我们不能准确及时地追踪海洋生态系统发生的变化。传统的海洋生物多样性评估主要侧重于大型生物,通过目视和人工方法进行,如潜水员调查、拖网、声学技术或航空图像。近期,利用遗传学方法进行的全球和局部地区的单细胞群落研究,揭示了海洋微生物和小型真核生物丰富的多样性。然而,特别是在相同的时间和空间尺度上研究分类群(如微生物到哺乳动物)和营养级的相互作用时,探索海洋中广泛存在的原核和真核生物的分布及其多样性仍然是一项重大挑战。 / v% z3 c2 ^0 u* z# s
在微生物生态学领域中,出现了环境样品的DNA扩增子测序(通常称为环境DNA (eDNA)宏条形码)方法,并且被应用于多细胞生物研究。尽管eDNA技术还未广泛应用于实践,但eDNA技术已经成为一种覆盖所有生物域的推测生态系统中生物共存模式的方法。目前为止,这些方法已经被用于研究横跨世界海洋中大部分区域的单细胞生物的空间分布和时间变异性。然而,该技术的另一个主要目标是全面探索完整的生物群落,调查单细胞和多细胞生物的多样性、分布以及环境参数,用以揭示群落之间的相互作用,并预测整个群落对环境变化是如何响应和应对的。 _6 v/ l5 Q: U3 ]! X* Q$ ^/ b
在此,我们分析eDNA宏条形码的时间序列数据,以评估海洋中不同分类群相对丰度比例的变化,并分析微生物到哺乳动物等生物之间的关联。这项覆盖所有生物群落、针对eDNA的多个保守遗传标记序列进行高通量测序的调查,阐明了群落动态和环境因素之间的关系。这种方法使我们能够检验不同分类群的丰富度和包括海面温度、叶绿素a和其他关键的环境和生物因素在内的多种环境变量之间的关联。通过网络分析,我们发现跨越不同营养级的共生生物群与环境参数的变化直接相关,这一研究结论明晰了群落对环境变化的潜在响应,并强调了共生生物和营养级间的相互关联。
3 r- ]9 m; D1 q* d9 x 2. 材料与方法 + U8 H2 F) V! T
在蒙特雷湾国家海洋保护区监测站点C1(36.797°N, 121.847°W),每两个月对“Rachel Carson”和“Western Flyer”进行一次采样。使用Qiagen DNeasy Blood&Tissue试剂盒对滤膜进行DNA提取。随后,对所有样品进行16S rRNA、18S rRNA、COI和12S rRNA基因宏编码。对来自四个文库(16S rRNA、18S rRNA、COI和12S rRNA)的结果进行生物信息学分析,得到物种注释结果。为了区分数据集内真正存在的OTUs与可能是假阳性的OTUs,我们对测序数据的占用率建模,然后进行测序数据的纯化。纯化后,进行测序数据的丰度索引,我们将所有的OTU按照它们的科注释进行分组,或者,如果一个OTU不能被注释为科,则被注释为目或纲。随之使用Kendall’s tau进行多重比较的相关性和校正;通过指定营养分配和附加的生活史特征进行分类群的营养分配;为了确定加权相关性,我们计算了包含所有样本中所有分类群之间所有成对Kendall tau相关性的相关矩阵后,为了将特定分类群与环境变量直接联系起来(图3),我们使用了R包mixOmics的稀疏偏最小二乘分析(sPLS)。 3 y! ]0 k) @3 D/ T4 J2 b
3. 结果与讨论 6 o# A2 G4 q. }. Q9 I% u
3.1. 群落时间结构
7 l7 ?0 B* y+ T 在美国Monterey湾国家海洋保护区(蒙特雷湾国家海洋保护区) 大约两月一次从长期监测站收集海水样品,并持续18个月(n = 8个时间点,2015年4月-2016年12月)。环境变量(包括水体的温度、盐度、溶解氧、叶绿素a和硝酸盐)均在样品采集时进行原位测量。用膜过滤器从海水中富集eDNA。同时,我们扩增了4个基因位点(16S核糖体RNA (rRNA)、18S rRNA、细胞色素c氧化酶I (COI)和12S rRNA)并测序。质量控制后保留了超过108条序列,最终鉴定出663个分类群。所鉴定的类群采用了不同的营养策略,包括腐养、自养、混合养和异养。由于扩增偏差可引起生物丰度和扩增子丰度之间的模糊关系,我们为每个注释的分类单元建立了从0到1的丰度指数。该方法假设扩增偏差来自于模板-引物的相互作用,并且对于任何给定的类群-引物对,这种相互作用在样本中都是恒定的,这允许我们推断不同类群之间的相对丰度变化。当一个分类单元被发现有多个遗传位点时,我们取这些指数的平均值来建立该分类单元的集合指数。然后,我们测量了类群eDNA指数的两两相关性(Kendalls tau),以检测同时发生扩增子指数丰度变化的类群。我们将“零模型”与多重比较结果进行组合,从而计算错误检出率(补充图3,修正的显著性阈值:tau= 0.70)。尽管一些陆生生物在其生命周期中具有水生阶段,并可能具有重要的生态意义,但鉴于我们对海洋生态系统的关注,所有出现在分类单元列表上的陆生生物都被删除了。除去陆生分类群后,还保留了348个独特的海洋分类群,其中274个(78%)在科水平上聚集在一起,其他则集中在更高的分类水平上(目、门等)。
- K- G$ P* B3 W* i4 |1 Q4 ]- M 为了评估群落比例丰度随时间的变化,对各类群的扩增子丰度指数进行了加权基因相关网络分析(WGCNA)。WGCNA建立了一个类群(节点)与其他类群(边缘)连接的网络,这些类群具有相似的趋势。相互连通的子网络(群落)是在主网络中确定的,代表了来自所有生命领域和营养级别的类群,它们的丰度随时间的比例变化具有很强的相关性。相关的变化可能来自直接或间接的营养相互作用,或来自对环境因素的一致反应。 , q( T) t9 U5 C
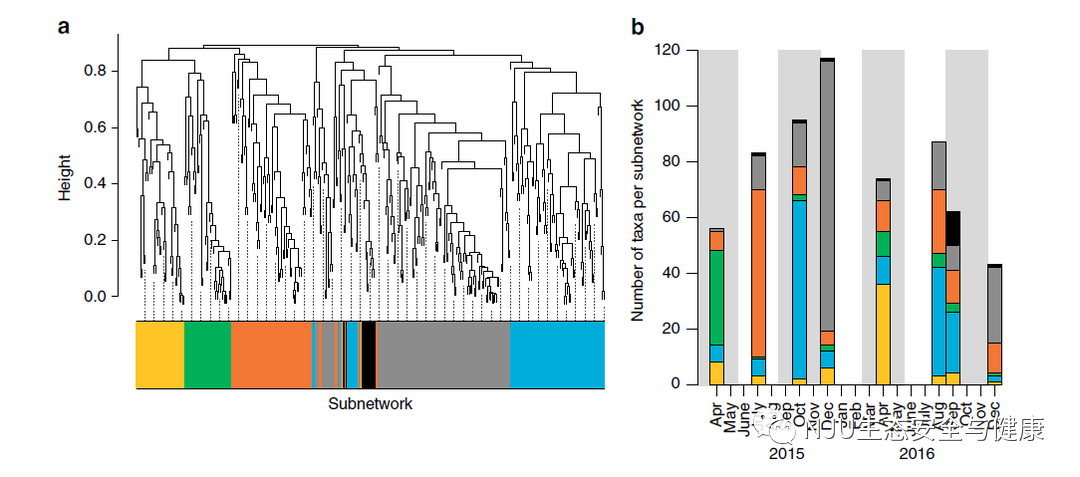
我们确定了6个子网(对于整个数据集的网络分析),代表不同的群落及其随时间变化(图1 a, b)。尽管从每个子网的代表类群主要集中在大多数时间点,但子网仍可以反映蒙特雷湾国家海洋保护区群落的季节性变化。对春季/夏季类群(橙色、黄色、绿色)和秋季/冬季类群(灰色、蓝色、黑色)的丰富度的eDNA观察结果明显不同(图1a、b)。所有的子网与环境变量相关:例如,类群丰富度的季节性(图1b)反映了冬季较低的叶绿素a水平(灰色亚网络,r = −0.76,Spearman相关p值= 0.03)和秋季较高的海表温度(蓝色亚网络,r = 0.74, Spearman相关p值=0.04)之间的显著相关性(补充图5和6)。
( [4 H7 J* y. o 2015年12月和2016年12月,在灰色网络占主导地位的情况下,类群的绝对丰富度(即总类群数)分别达到最高和最低。子网内多样性的升高/降低,表明了群落是如何应对年际变化的,例如,在2015年12月由于强烈的厄尔尼诺事件引起平均海面温度和物质循环变化(补充图7),这在之前已经有eDNA的研究观察到。该地区的其他研究也报道了厄尔尼诺事件期间群落结构相对于其他年份的变化,包括当分类群从暖和的南部或近海岸往北转移至加利福尼亚海岸中部时物种丰富度的异常变化。 0 A$ S W3 O* |7 {, I% a# ?
% H1 c9 i: l' G0 b1 ~
图1 eDNA数据与环境变量相关的加权基因相关网络分析 7 c* u4 ^0 M$ ^) v
a. 基于所有分类群丰度指数变化的聚类分析,使用Kendall相关系数的树状图。这些颜色对应于不同的子网。b. 结合图1-a,在采样时间点上,每个子网中观察到的分类群丰富度显示了2015年12月的最高累积丰富度。阴影部分代表春天和秋天,白色代表夏天和冬天。 4 o S" M+ @, Y' Q
3.2. 子网络的组成和时间行为 . |+ Q0 C. i: C6 S
具有强营养相互作用的类群在时间上具有高度相关的扩增子丰度模式。为了评估这些营养级的相互作用,我们假设除了外共生和内共生的生物之外,不同营养级别的类群之间的相互关系不是直接的。我们观察到在类群之间有一些可能的间接联系,以及表明营养相互作用的各种直接联系。占主导地位的2015年和2016年的春季子网(分别为绿色和黄色)包含的大多数中心硅藻类群(分别为30%和41%)最高比例的丰度(补充图8)。这些子网络与早期的季节性上升流有关,自养群落的多样性最低,尽管已知中心硅藻会在春季大量繁殖。在蒙特雷湾国家海洋保护区中,典型的羽状硅藻出现在中心硅藻之后;然而,我们发现羽状硅藻物种丰富度在所有子网(以及时间)中分布均匀,2015年春季和冬季的丰度比例更高(补充图8)。夏季/秋季子网(橙色和蓝色)包含了大部分甲藻类群(分别为26%和20.5%)。预计这将发生在上升流季节的后半段以及过渡到非上升流状态。然而,在我们采样期间,灰色子网络(在2015年12月和2016年12月占主导地位)含有最高比例的甲藻(35%),这与当时最小的季节性涌升相一致,并在2015年底因厄尔尼诺条件而加剧,当时冬季海面温度可能高于平均水平,更容易分层。除甲藻丰度较高外,桡足类丰度比例在2015年冬季也最高(补充图8),占当时总类群数的38%(灰色子网)。 + I3 u `3 s1 w3 H% O
秋季(蓝色)和冬季(灰色)子网络与环境参数的相关性最强。我们重点关注这些子网,以说明环境条件和假定的生态响应之间的关系(补充图5和9)。秋季(蓝色)的优势子网由81个分类群组成,当海面温度> 14℃时更常见(图2a-c和补充图7)。蓝色子网包含可导致有害藻类大量繁殖的浮游植物分类群(例如,有毒藻类科Gonyaulacaceae和Prorocentraceae),之前已观察到这是在蒙特雷湾国家海洋保护区中正温度异常的反应。Balaenopteridae(序列鉴定为座头鲸)在蒙特雷湾国家海洋保护区中非常常见,其是一种顶级掠食者,也是蓝色子网中联系最紧密的分类单元(最中心的节点,见方法)(图2a)。磷虾(磷虾科)是座头鲸的常见猎物,被归入同一子网,但其与鳞虾科的相关性不强(图2,补充图10和11)。 * z8 j3 O" W! o- P
优势冬季(灰色)子网由114个分类群组成,与低浓度的叶绿素a正相关(图2d–f)。在这个子网络中,我们观察到在同时具有高扩增子丰度指数的分类群之间可能存在直接联系,即小樽科(序列被确定为加利福尼亚海狮;一种顶级掠食者)和鲹科(序列确定为太平洋竹荚鱼,次高营养级)。加利福尼亚海狮捕食包括太平洋竹荚鱼在内的杰克鱼,这种鱼出现在蒙特雷湾国家海洋保护区中。杰克鱼经常以浮游桡足类为食,事实上,灰色网络中的桡足类(尤其是 Metridinidae)与杰克鱼密切相关(r=0.80,Spearman相关p值< 0.05)。灰色子网包含几个具有寄生生活史的分类群,包括共甲藻科(甲藻属),它们寄生于广泛的宿主;包括甲壳动物、放射虫和鱼类,它们也存在于灰色子网中(补充图8、10和11)。大多数放射虫类(90%),包括所有Acantharea形态,与2015年冬季(灰色)子网有关。硅质(多囊藻)和放射虫可以吃其他浮游生物,一些分类群有沟鞭藻、单胞藻或共生藻内共生生物,如冬季网络中所见,这可能使这一群体的成员在硅藻不太普遍分布的时期占主导地位。因此,eDNA代谢与网络分析相结合的方法解释了共生分类群的子网,我们可以用它来发现假定的生态相互作用。 9 J: I+ C h8 E
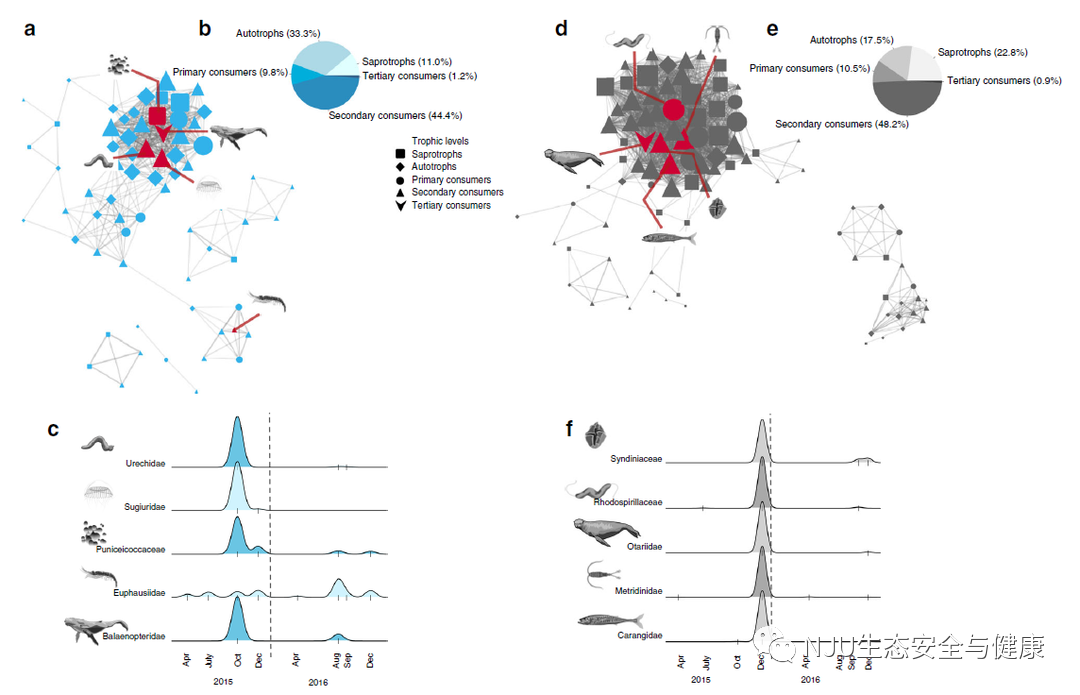
6 _0 G; {! n* {" F. g 图2 特定类群随时间变化的网络和扩增子指数丰度
, E2 i3 C. f% w) I% T 2015年和2016年秋季(蓝色)子网(a-c)和2015年12月(灰色)子网(d-f)的网络可视化。蓝色(a)和灰色(d)子网以节点(分类群)和边缘(相关性)显示,代表各个分类群之间的联系。为了最好地显示子网,仅显示网络连接(边权重)高于阈值0.2的分类群(即,去除了低相关性,参见边计算方法)。在子网内检测到的不同营养级用不同的符号表示,参见图例。这些分类群说明了蓝色和灰色子网络中共存分类群的复杂性,在此基础上选择分类群(c)和(f)的分类群表示。(b)和(e)表示每个子网内营养级的分布。分类单元节点符号的大小与单个分类单元在子网内的边连接数量有关,边的宽度代表两个给定节点之间相关性的权重。每个子网内的红线指向子网内高度相关的分类群(r > 0.9)或高度关联的分类群(前10%以内),在(c)和(f)中突出显示。在我们的采样期间,这些分类群的比例丰度在(c)和(f)中给出。图(c)和(f)x轴上的刻度线代表我们对这些分类群进行观察时的采样次数,y轴的高度是相对于每个分类群的观察次数而言的,并不代表绝对丰度值或生物量。垂直虚线表示2016年1月1日。
1 f2 H @' u2 F! P 3.3. 网络与环境变化的对应关系
; T- [, ]* R4 \& r% t+ f. y 蓝色(秋季)和灰色(冬季)子网分别与高温和低叶绿素a浓度相关联,表明生物群落的连贯转移与环境条件的变化相关联。在这些子网络中,组成分类群的扩增子丰度指数(见方法)可预测地与相同的协变量相关(图3a,例如,来自冬季子网络的大多数分类群与叶绿素a负相关,也与溶解氧负相关)。
# L' G, Q7 x2 Y( i5 D 在秋季子网中,大多数与温度正相关最强的分类群也是具有最高子网内连通性(见方法)和最高子网要素关系(即子网与其他分类群的相关性(或边缘权重)之和最高)的分类群(补充表3)。对于秋季和冬季子网,单个分类群与环境变量(分别为温度和叶绿素a)的相关性(Spearman r相关系数)与这些分类群的关联度显著相关(即,分类群的关联度越大,与环境变量的相关性就越显著,图3b,c)。 3 ~0 x, \- ]6 x5 n0 ^1 q/ N- o
最近的研究表明,在微生物研究中,中心性的程度可以以85%的准确率识别关键分类群,尽管这也遭到了一些批评。这种预测能力尚未在从微生物到哺乳动物的跨营养级研究中得到检验。在最相关的分类群(前10%)和与温度或叶绿素a显著相关的分类群(图3b,c,红色虚线上方)中,我们发现了所有营养级和所有生命领域的代表。从秋季(蓝色)子网来看,不同营养群的顶级代表包括棘螠科(多毛纲)、芮氏水母科(原生动物)和浮霉状菌目(细菌)(补充表3)。这些分类群分别代表次级消费者、初级消费者和腐生物。
1 J, ]0 m9 f. x8 z1 Z, l 在冬季(灰色)子网络中,来自前10%相关分类群的代表是放射虫纲(放射虫)、鲹科(杰克鱼)和Thalassoarchaea (古细菌),分别代表一级消费者、二级消费者和混合消费者。每个子网络中的第三级消费者(座头鲸和加利福尼亚海狮)也分别属于排名第一和第四的10%最高度关联的分类群(图3和补充表3)。 . Q# h* E& ^4 X8 |* ]
与各自子网内的其他分类群联系最紧密的单一中心分类群是蓝色子网的鲸豚类(座头鲸)和灰色子网的红螺菌科(变形菌)。这些分类群在其各自的子网内共连接到24个(29%)和48个(42%)分类群。两个子网络中排名最高的分类单元之间连接的分类单元数量的差异是由于该特定子网络中分类单元数量的差异(蓝色:n = 81,灰色:n = 114)。
% [4 P4 U! q `" v) y; F7 w 有一个可量化的阈值来对指标或重点分类群进行一致的识别和验证是很重要的,并且需要实验证据来显示各个分类群对群落功能和组成的影响。仅网络相互作用就能显示出不太可能直接相互作用的分类群之间的积极联系(例如,棘螠科(一种沉积物栖息动物)和座头鲸),重要的是要记住,这种相关性显然不是因果关系。然而,这种假定的相互作用网络突出了以前可能没有注意到的分类群与其环境的波动,因此可能是识别新生物和群落组合的强有力的方法,可以指示环境变化和先锋保护策略,这将特别受益于物种水平上对这些类型的分析。
; Q% o: W3 x8 v 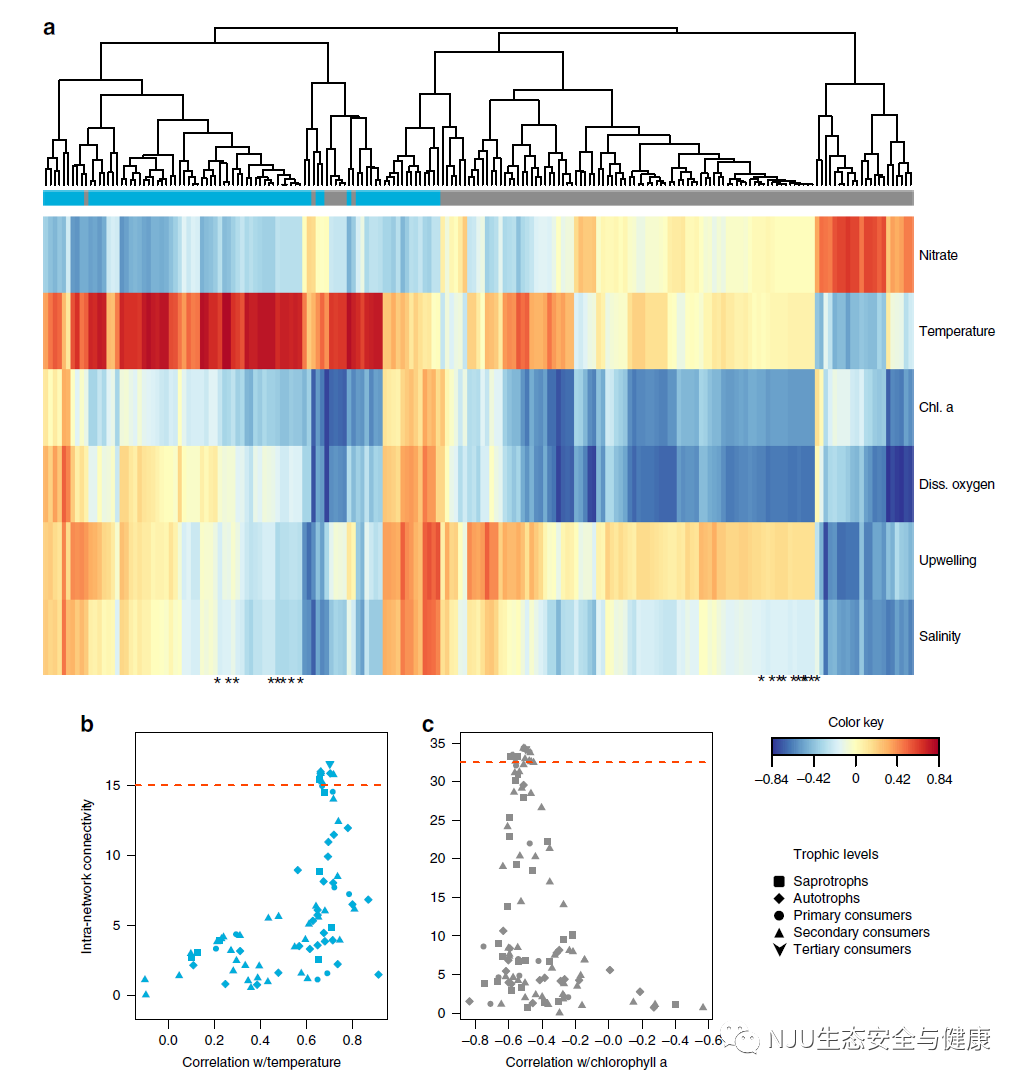
( C/ _9 t' u1 |% |
图3 分类群和环境变量之间的偏最小二乘分析,以及生物类群与环境的连通性和相关性图 8 V8 W$ y6 c/ @/ H8 N4 C% h0 L
(a)显示灰色(冬季)和蓝色(秋季)子网(叶色)中所有分类群(列)与环境变量(行)相关性的偏最小二乘分析图,通过Kendall相关系数进行聚类。热图由相关系数值着色,参见颜色键。星号代表每个子网中前10%最相关的分类群(在(b)和(c)中红色水平线的上方)。蓝色(b)和灰色(c)子网内每个分类单元的连通性或中心性的相关性分别与温度和叶绿素a相关。符号对应于图例中显示的营养水平。 5 ~2 K& R4 [8 ?( {2 E
注:“材料与方法”、“致谢”、“补充材料”、“讨论”、“参考文献”等部分内容未放入推送中,可至文献原件中查看。
4 _+ j/ {. U4 r; Q& V3 \5 N u (为使文章结构更具可读性,对部分行文结构进行了重构)
. W6 @1 S8 P! P1 w' Q/ t 翻译| 钟文军 黄湘云
/ }0 d' M" U- _- g G1 { 校对 | 张颜 姜山
! v2 y/ C) q' k) n/ m3 V$ W 来源 | 生态毒理与健康风险研究团队——河流生态组  6 U7 L8 `- H5 z5 {2 F% I) q
6 U7 L8 `- H5 z5 {2 F% I) q 长按识别上方二维码 + J, q$ X0 t' U$ l
关注我们 ! V9 T+ M# M5 v
THE END
+ {* w* @1 z: P9 p- g- g. _9 O; t/ W8 x' g# ?% g
5 ^* W* ^ M, C& k
* M/ Q& x! Z b- |% f
0 ?2 a6 b# E/ f6 K | 

