|
) Y3 T+ f' ~# c3 Y6 j
一 海洋环境梯度 8 {7 I, Q7 g0 a4 q5 Y
——什么是海洋环境? 2 i8 i- L" U# j, q: B# K3 M
1.总体特点:具有复杂的时间空间性质,及理化生物性质 2 |" t: p$ N' q; ~7 y% f
2.海洋以两大动力系统(风生/热盐)和丰富的生命活动驱动了物质和元素的生物地球化学循环,塑造了异常丰富的海洋环境区域和微区 7 O, G% r6 Z$ v4 b. h( v. S" r
3.海洋环境界面:海-气、海-陆、海-泥、水-生 " P& D$ Q4 O: p7 @' X4 U: i
4.特殊的海洋环境:河口、海底热液/冷泉、冰川、酸化、海洋沙漠、水团等, 3 @1 }* J" O. S0 F7 m, h
形成于理化性质变化剧烈的界面区 8 o9 [& n" {. _/ p
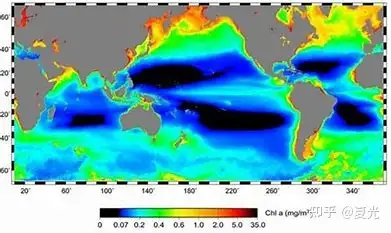
5.海洋环境三大梯度:①纬度地带性。随纬度变化的水平梯度;②环陆地带性。水平方向上,从沿海向开阔大洋的梯度;③垂直梯度。随深度变化的梯度[1]。 * F0 k: k5 u; ~7 C; ]6 z7 J
图一 海洋初级生产力的纬度地带性分布二 海洋环境纬度地带性
* E1 Q5 T) F; t: E% K 1. 概念:指气候、水文、生物、底质等自然要素及自然带沿纬线方向带状分布,并沿纬度变化方向逐渐更替的现象。
0 Y' q( n4 P: M0 g4 D 2. 分带:从赤道到高纬度极地,海洋环境被分为热带、亚热带、温带、亚寒带和寒带不同的热量带。
: H% h' u0 U- T" h4 c) c/ [) V 3. 成因:太阳直射角的纬度地带性变化,造成海洋热量分布呈现明显的纬向带状分布;季节变化。 " Q) `& B* H) Y, v! Q& x
4. 实例: 7 Y6 ~5 V2 X {8 C
(一)南海大气温室效应的纬向地带性
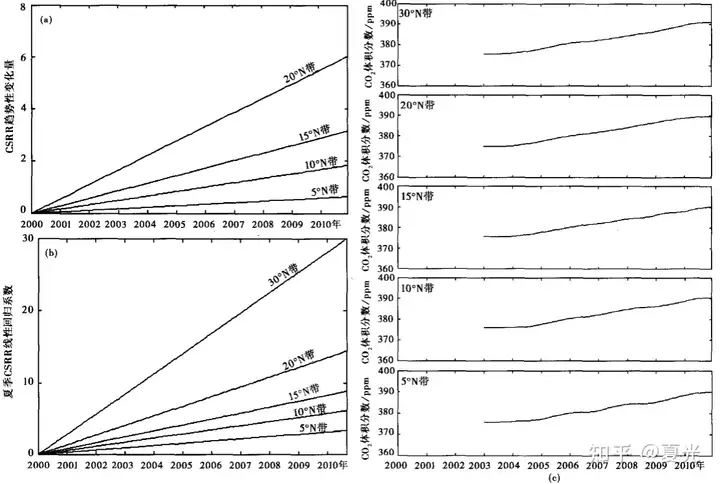
. r0 X3 z: ^5 w( @ 南海温室大气的主要分量为由低纬度往中纬度逐渐增加的,具有明显纬向地带性的水汽大储库和纬向地带性较弱的CO2小储库长期联合影响[2]。 + a* a1 t0 P, z3 ]: {+ _4 y
受CO2等温室气体影响,东亚海域温室效应呈上升趋势,其变化量具有典型纬向地带性分布特征,低纬度海域为温室效应低增长区,中纬度海域温室效应增大。 & C) Q6 |& \+ e1 t" {
图二 南海各纬度带的CSRR趋势性变化量(a),以及南海与东海各纬度带的夏季线性回归分析结果(b,已移除偏移量)和CO2趋势项分布(c)(二)上层海洋浮游生物地理分布
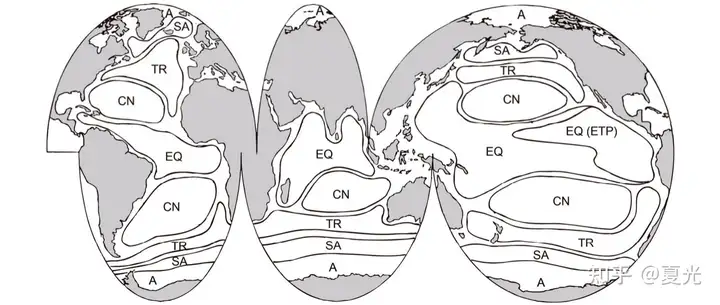
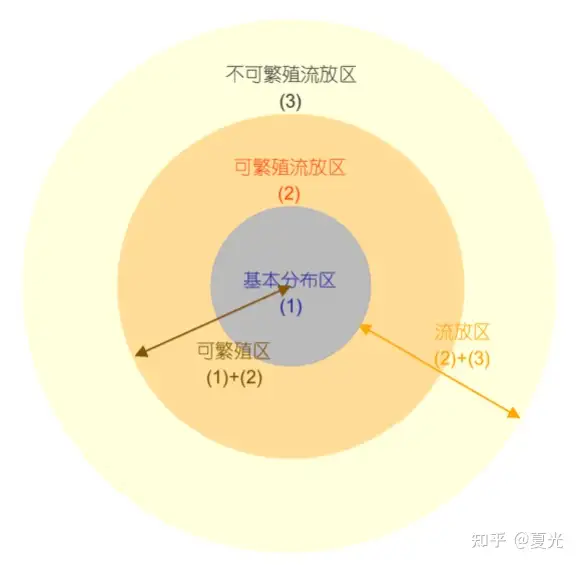
/ T4 _5 m! d$ a 大洋浮游生物的地理分布特点呈纬度平行的九条区带,分别是:赤道条带、2个中心区条带、2个亚极区条带、位于中心区和亚极区之间的2个过渡区条带,及位于南北极海区的2个极区条带。
' g/ o% g% \3 i 洋流和水团是生物分布特征的决定因素。过渡区条带的可能调控机制为中尺度涡。相同分布格局的生物扩散能力不同,扩散能力最差的生物是核心种,最强的为优势种[3]。
6 s. i% P3 j" C- s" l; Y* q8 c( D) D 图三 海洋浮游生物地理分布的九带式分布格局[改绘自Longhurst(2001)]图四 浮游生物分布区的空间结构示意图(三)印度洋长鳍金枪鱼(Thunnus alalunga)时空分布 ' {, X @; j+ c7 v
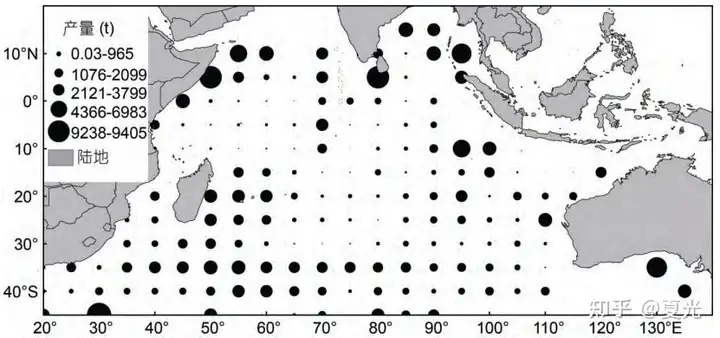
海表温度直接或间接地影响渔场的分布、洄游的路线等。海表盐度贡献率低,对长鳍金枪鱼分布影响较小,其最适宜范围在 34.5—35.5‰。叶绿素 a 浓度通常影响长鳍金枪鱼的食物链,从而间接影响长鳍金枪鱼渔场的分布[4]。 . J2 R4 n# O' W
图五 印度洋长鳍金枪鱼2012-2019年累计产量分布图海洋环境纬度地带性的生态效应总结: ' D( e) _ S, S: P- `, p
① 海洋环境的纬度地带性是多对象多要素的;
; x8 R! t J! ]+ m* [& M2 w ② 海洋环境要素的纬度地带性分布(温度、盐度、叶绿素浓度等)受到日光等自然条件和海洋生物共同影响,生物能实现跨纬度分布;
* X* s7 R0 u" }% m1 n5 J ③ 生物的分布和多样性对环境要素的纬度分布具有一定的适应性和抵抗性; - v4 c; B$ D2 |: w" S
④ 海洋生态响应是多要素相互耦合的。 $ V9 z& E2 E; ^5 f; T6 H
三 海洋环境环陆地带性
! q' ]5 ^- U9 a. r! t; } 1. 概念:指气候、水文、生物和土壤等自然要素以及自然带从沿海向内陆逐渐更替的分布规律。也就是主要因水分条件的变化所致,又称“干湿度地带性”。 % S. p) e* O) ^5 Q4 w n2 a0 ?
2. 分带:根据距离陆地的远近,可以分为河口、滨海、浅海和开阔大洋。 3 E: F1 o# q5 |2 F: s& c
3. 成因:距离陆地的远近,造成海陆交换性质的不同;海流性质以及营养盐含量的差异;温度和盐度差异 & c M& f, F0 C' o9 ~" T
4. 实例:——南流江河口桐花树生物动力地貌过程研究 ! X4 Q: ^0 t( ^& W6 [
南流江河口桐花树自西北向东南快速向海扩张的格局与潮滩向海淤积前展维持一致,潮滩滩位升高成为宜林滩地是桐花树向海扩张的基础;桐花树种群的消浪作用减缓水动力强度,促进泥沙在潮滩沉积,由此加速潮滩发育;低潮滩淤涨为中潮滩、高潮滩,为桐花树种群扩张提供立地条件,促进种群发育扩张[5]。 , s5 w- z" k' a- \7 n3 |2 [
5. 海洋环境环陆地带性的生态效应总结: 2 e) z/ ]4 o4 n4 v3 ?/ Q
①复杂的物理化学成因——河口动力学导致生物类群丰富多样; ; G# k( L1 ?3 v7 k( Z) I
②河口地区频繁的人类活动造成的污染问题显著,以毒素降解、生物修复、环境友好的生物类群优势;
0 n# p3 ?; ?# W. | ③陆海交接处存在广温、广盐的生物适应机制。
! q, q! a! [3 P* @: r- v% m, g' g2 _% _5 B 四 海洋环境垂直梯度 $ A# K: j( Y' @. T/ X; h
1. 概念:指随海洋深度变化的理化要素的垂直梯度现象。
' H. e* z. v& O; b9 T 2. 分带:海水水温的垂直分布可分三层:①混合层,一般在大洋表层100米以内,由于对流和风浪引起海水的强烈混合,水温均匀,垂直梯度小。 ) z& A ^% h/ D. B3 U( Q( Q% D( Y) Z6 Y
②温跃层,在混合层以下和恒温层以上,水温随深度增加而急剧降低,水温垂直梯度大。
# v. U# ]! y; W6 l ③恒温层。大洋深层水体,温度主要来源于上层水体热量交换,伴随海洋底质的热量输送,温度扰动小。 % p9 e1 K4 `. X B; f) W+ Y* a
3. 成因:海洋物理空间的深度变化;温度、盐度和压力等环境要素的变化 . J& i3 P3 J j ^) ^
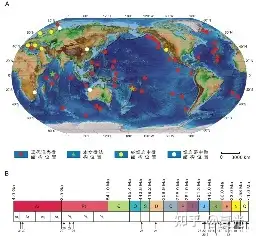
4. 实例:——海底玄武岩玻璃中蚀变微结构
4 B+ B0 l! U9 {# s# R. }% ] 玄武岩玻璃中的生物蚀变微结构为微生物摄取玄武岩玻璃中营养成分,通过新陈代谢产生有机酸溶解玄武岩玻璃而形成的微米级孔洞或钻穴。生物蚀变微结构在现代海洋洋壳及代表古老洋壳残片的蛇绿岩和绿岩带中广泛存在[6]。
5 A) _/ N- z# S V9 K8 U) G7 r 图六 全球海底玄武岩玻璃形态及分布图5. 海洋环境垂直地带性的生态效应总结:①跃层的存在,导致在海洋垂直分层地带产生丰富的生物分带性;
5 I( ]; D" P+ N" @! f ②生物活动受到海流的动力学因素影响(热盐环流和风生环流),存在温度、DO、营养盐、光照、压力等显著变化要素影响等。 5 m; y) i3 C8 p( ?! ^
③海洋表层的初级生产和海底的初级生产途径差异大,造成海表和海底的生态系统差距大。
$ x. u% o' ]( N7 u$ M 参考文献 " Q' Q$ v& N B+ ~- b
[1] 张庆忠,陈庆义,吴文良.景观生态学:海洋生态系统研究的一个新视角.生态学报,2004,24(4):819-825.
; Q3 b- S; g! ]6 Q' s, U; y [2] 陈嘉伟,李炎,邵浩,等.南海大气温室效应纬向地带性的遥感观测[J].第四纪研究,2016,36(1):131-143. ' r& H1 x- L3 ?- f' u6 o
[3] 张武昌,赵苑,董逸,等.上层海洋浮游生物地理分布[J].海洋与湖沼,2021,52(02):332-345.
; O1 R+ s# h3 v8 y3 O [4] 程懿麒,张俊波,汪金涛,雷林.基于神经网络的印度洋长鳍金枪鱼(Thunnus alalunga)时空分布与海洋环境关系研究[J].海洋与湖沼,2021,52(04):960-970+1-6. ) q* n0 i& T n
[5] 王日明,戴志军,黄鹄,等.南流江河口桐花树生物动力地貌过程研究[J].海洋学报,2021,43(09):102-114. % |8 o$ P- p9 {, V" q8 @/ A6 C
[6] 陆炫臣,颉炜,罗茂,等.海底玄武岩玻璃中蚀变微结构:探索海洋深地生物圈的新材料[J].古生物学报,2021,60(02):281-298. ( Q5 o, }3 m; c5 W
/ P: i; ~" [7 k% Q2 i$ m3 X& I
. G4 _) x; J1 j2 L
. p7 N% l/ R6 P3 u, g8 x- t/ n$ D$ }8 _# U6 \: O0 P, R
|