( f, W. ^ v$ F# C9 |0 t: V6 Z- ~ 把一篇本科课程作业改改发上来吧,但也没加今年的新成果。这领域成果更新还挺快的,我这玩意不发出来过几年也没用了。希望不会有什么无良中文期刊抄到这篇知乎回答上来 _(:з」∠)_
0 E. m/ Y0 N& l& Z6 J( N. d% V9 s
在全球变化的背景中,CO2上升可以同时引发海洋酸化、气候变暖和海平面上升,这些变化都可能对珊瑚产生负面影响,而珊瑚对这些变化的响应是非常复杂的。本文就简单介绍一下海洋酸化、气候变暖和海平面上升下珊瑚以及珊瑚礁系统可能的响应。
7 z. |" N% i6 c( d( [8 `! i0 b( k 1 海洋酸化下的珊瑚礁
6 c; n' [1 w. p 随着大气CO2浓度的上升,海水pH正在发生下降,CO32-的浓度也会发生下降,这会使海水碳酸钙饱和度下降,从而影响造礁珊瑚的钙化情况(reviewed by Hoegh-Guldberg et al., 2007; Pandolfi et al., 2011; but also see Hughes et al., 2017a)。
0 D, o9 U1 e& ^ 1.1 海水酸化对珊瑚钙化的影响机制
& j/ W4 ?7 s4 n. }& I% `7 y
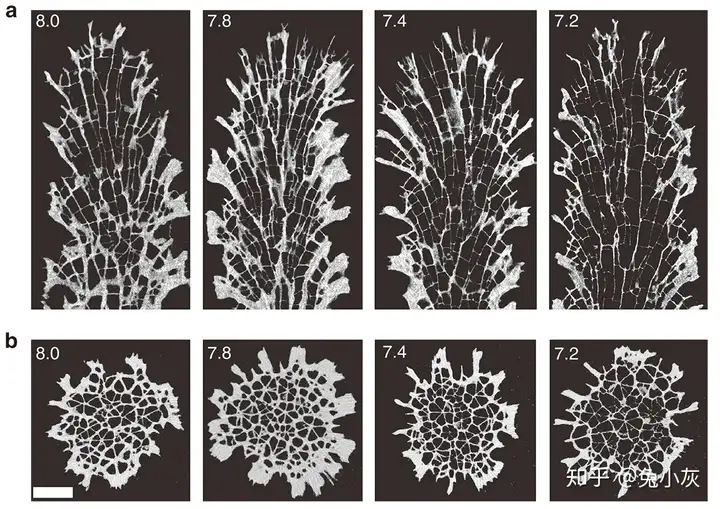
珊瑚的钙化率由珊瑚的骨骼密度与生长率两项参数决定,与直觉不同,实验室(Tambutté et al., 2015)和野外研究(Crook et al., 2013; Fantazzini et al., 2015; Mollica et al., 2018)都表明酸化并不会显著影响珊瑚骨骼的生长率,而是使骨骼密度发生了显著下降,具体而言,是使骨骼的孔隙度发生了增加。并且扫描电子显微镜(SEM)和原子力显微镜(AFM)的观察表明不同pH下生长的珊瑚的文石晶体结构没有明显区别,也没有溶解的迹象;但显微CT(micro-CT)的观察发现珊瑚骨骼的形态结构发生了变化,即骨骼变薄,珊瑚体占的空间变大。这说明酸化下珊瑚钙化率的变化不是由于简单的物理化学溶解过程导致的,而是由珊瑚的发育程序控制的。
8 n2 F: U/ B' w 酸化条件下珊瑚骨骼孔隙度增加,导致钙化率下降(引自Tambutté et al., 2015)对于幼年珊瑚,酸化条件下不但骨骼的形态结构发生了改变,骨骼表面也表现出了溶解的迹象,说明幼年珊瑚可能对于海洋酸化更脆弱(Foster et al., 2016)。
* c# s9 J" W+ }" P$ H7 V
1.2 珊瑚钙化对海水酸化具有抵抗力
3 e8 T, D7 b, f* B. D- F4 e9 e
虽然海水酸化会导致环境海水中的文石饱和度下降,并确实会影响珊瑚的钙化率,但珊瑚还是存在一定生物学机制来减轻酸化对生长的影响。
' L: w- }* q6 f" E! G* ]% V8 z8 p' {% X 珊瑚的骨骼钙化过程并不直接发生在环境海水中,而是发生在珊瑚内的胞外钙化流体(extracellular calcifying fluid)中,其与腔肠中的海水被反口面内胚层、中胶层和外胚层分隔开(Al-Horani et al., 2003)。多种不同的实验方法都表明珊瑚钙化流体中的pH要高于环境海水的pH,其中原位的微电极和pH敏感染料测得的较瞬时的日间钙化流体pH增高约为0.6-1.2(Ries, 2011; Venn et al., 2011),而硼同位素测得的较长期平均的pH增高约为0.3-0.6(McCulloch et al., 2012)。并且当海水pH下降时,钙化流体中的pH下降幅度要小于海水pH的下降幅度(McCulloch et al., 2012),从而将骨骼钙化实际发生处的文石饱和度还维持在相对较高的水平。珊瑚细胞膜上的由ATP驱动的Ca2+/H+反向转运泵应当参与了这一pH调节过程(Al-Horani et al., 2003)。理论计算(McCulloch et al., 2012)和实际观察(McCulloch et al., 2017)都表明pH上调过程消耗的能量相对不大,应该不太影响珊瑚的生长。
$ j! u6 A k" ~. ^1 B% h2 z McCulloch et al.(2012)考虑了珊瑚上调pH的能力,建立了IpHRAC模型以模拟全球变化下的珊瑚钙化情况。其结果表明若假定温度是恒定不变的,CO2增加引起的海水酸化还是会导致珊瑚的钙化率有一定下降,但下降程度要比其他没有pH调节能力的钙化生物小一半以上;而若同时考虑CO2增加引起的变暖效应,珊瑚的钙化基本就不会发生变化,对于某些pH调节能力较强的种类钙化率甚至还可有小幅上升。
/ m7 P, z @6 n( C9 S2 X: N" [+ B 对自然环境中澳大利亚西部珊瑚礁的研究表明,1900-2010年间,滨珊瑚(Porites)的钙化率整体上也确实没有下降,并有一定上升趋势;整体上海表温度距平与钙化率距平存在正相关关系(Cooper et al., 2012)。这表明至少当温度增加不太大时,又考虑到珊瑚自身对海洋酸化的抵抗力,全球变暖可以抵消海洋酸化对珊瑚钙化的影响。
% y% ? k: M+ A1 y: Y
1.3 海洋酸化可能抑制珊瑚幼虫的定殖
3 b8 \5 {4 R" q$ r9 T2 o! F 尽管珊瑚的顶端生长及整体的钙化率对海洋酸化可能不太敏感,但珊瑚的侧向生长对酸化却较为敏感(Holcomb et al., 2014)。侧向生长位置的钙化流体pH调节能力要相对较弱,这可能会影响珊瑚向新的基底扩展的过程,而这可能对于珊瑚幼虫或碎片在新基底上定殖的过程尤为重要。因此在评估海洋酸化的影响时,不能仅考虑其对于整体钙化率的影响,而是还要特别注意考虑这些对酸化敏感的生长阶段。
( K1 @' U# E! n4 S 另外,壳状珊瑚藻是珊瑚幼虫重要的募集附着基质(Heyward & Negri, 1999),但由于壳状珊瑚藻含有较多的高镁方解石,其对于海洋酸化非常敏感(Morse et al., 2006; Kuffner et al., 2007; but see Vásquez-Elizondo & Enríquez, 2016; Cornwall et al.,2017, 2018)。实验也确实观察到了海水酸化会导致壳状珊瑚藻相关的微生物群发生改变,并抑制珊瑚幼虫在珊瑚藻上附着(Webster et al., 2012)。这表明海洋酸化可能会特别对受损珊瑚礁的重新恢复产生负面影响。
x g8 |' F& T. K& J B3 A3 o/ [ 1.4 海洋酸化可能引发珊瑚礁的溶解
# ~! J5 b5 O- d* N1 x5 R3 F
虽然活体珊瑚对海洋酸化有较强的抵抗能力,但自然环境中的珊瑚礁并不仅有活体珊瑚,礁沉积物也是珊瑚礁的重要组成部分,其包括了死亡珊瑚的骨骼、其他生物的沉积,以及各种无机沉积。显然海洋酸化对珊瑚礁的影响不仅与活珊瑚的钙化有关,同样也与礁沉积物的溶解有关(Eyre et al., 2014)。
; E- E" W0 G; P; P2 G @5 ~4 A 活体珊瑚可以通过上调pH抵抗海洋酸化,但容易看出,珊瑚礁中没有生命的部分显然无法主动对pH进行调节。对于在酸化水体中生长7个月至一年以上的活珊瑚,其文石晶体的形态没有明显变化(Rodolfo-Metalpa et al., 2011; Tambutté et al., 2015);但对于暴露在外的死亡珊瑚的骨骼,在酸化水体中放置3个月文石晶体的形态就已经有明显变化(Rodolfo-Metalpa et al., 2011),说明其发生了较明显的溶解,这验证了死亡的珊瑚骨骼对于海洋酸化比活珊瑚敏感的多。
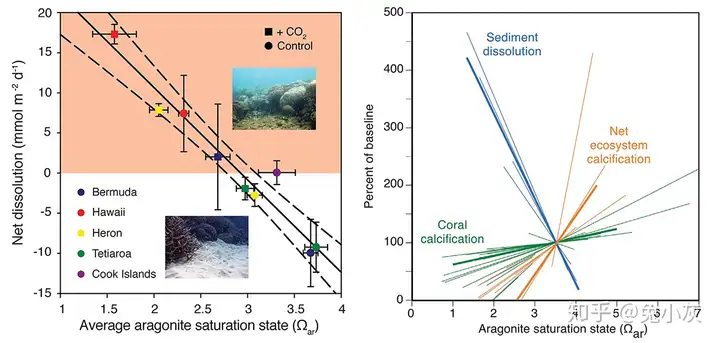
4 j1 A9 y4 Y$ |9 a- I Eyre et al.(2018)及Cyronak & Eyre(2016)在太平洋和大西洋的多个珊瑚礁中,通过向置于礁沉积物上的有机玻璃腔通入CO2的方法控制局部海水的酸度,表明沉积物的溶解与海水的文石饱和度有显著的负相关。对于夏威夷的珊瑚礁,自然情况下礁沉积物就已经处于了净溶解的状态。虽然总体上目前礁沉积物还处于净沉积的状态,但按照模型,目前的沉积相比工业革命前已有下降,并且在本世纪中期就可能会转向净溶解,随后整个生态系统的净钙化也会变为负值,这可能会导致整个珊瑚礁生境的破坏。
1 t# q/ h- Y: g2 Y5 Z3 K- j
礁体碳酸钙的溶解比珊瑚钙化对海洋酸化更敏感(引自Eyre et al., 2018)Albright et al.(2016)通过向自然珊瑚礁中的泻湖海水加入NaOH以调节海水的碱度,结果表明若将海水pH值恢复至接近工业革命前的水平,整个珊瑚礁系统的净钙化率会有显著上升。这或许表明通过地球工程的方法增加珊瑚礁周围海水的碱度是一种可能的珊瑚礁保护措施。
: v2 M4 a8 a& w6 w 1.5 海洋酸化会改变珊瑚礁系统的群落结构
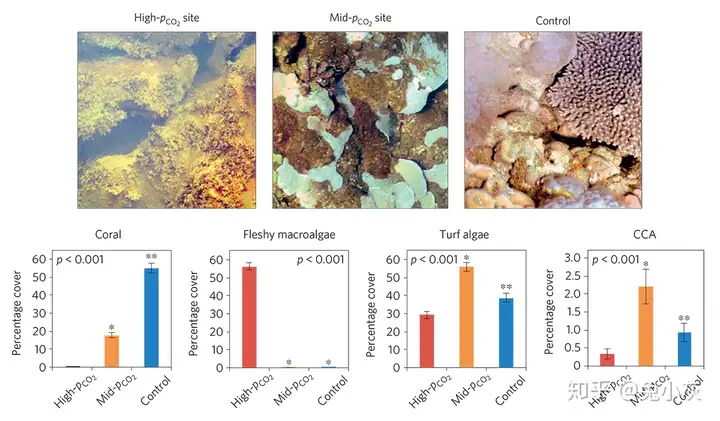
1 m. i5 M l' T, ^+ v f 已经有多项研究关注了与海底火山活动相关的CO2喷口或渗漏点附近的天然珊瑚礁群落。这些研究表明,在空间上随着pH由高向低转变,生态系统可能会由珊瑚主导转变为大型藻类主导(Enochs et al., 2015),珊瑚群落可能会由造礁的硬珊瑚转变为非造礁的软珊瑚(Inoue et al., 2013),珊瑚的多样性可能会发生减少(Fabricius et al., 2011),礁体的发育可能会停止(Fabricius et al., 2011),对珊瑚的生物侵蚀可能会发生明显增强(Barkley et al., 2015)。另外与珊瑚礁相关的大型底栖无脊椎动物(Kroeker et al., 2011; Fabricius et al., 2013)以及下层浮游动物(Smith et al., 2016)的数量以及多样性总体上也会减小。尽管这些研究直接观察到的是与空间pH变化相关的群落变化现象,并不是在时间上pH变化后带来的影响,但其仍然强烈暗示了全球海洋酸化也可能导致类似的珊瑚礁系统的群落结构变化。
' r, V) V3 T; o! G 酸化可能会使生态系统由珊瑚主导转变为大型藻类主导(引自Enochs et al., 2015)另外实验室研究还表明海水酸化可能会引发与珊瑚共生的微生物群落结构发生改变,这可能会改变珊瑚的生理或健康状况(Meron et al., 2011)。
" I2 }% {/ h8 M6 }
2 全球变暖下的珊瑚礁
8 B* [" p, y" | P! X& D+ s
随着全球气候系统的变暖,热带海洋的温度也在上升,而温度上升可能会使造礁珊瑚丢失其共生藻,即为珊瑚白化现象,若热胁迫持续存在,则白化的珊瑚最终可能会面临死亡,对珊瑚礁生态系统造成严重破坏(reviewed by Hoegh-Guldberg et al., 2007; Pandolfi et al., 2011; Hughes et al., 2017a)。
: @5 Y7 F T3 G" f
2.1 高温胁迫引发珊瑚白化的生物学机制
$ k. ]% J9 G' K 对于珊瑚白化在分子以及细胞层面上的机制,目前研究的还不很充分。有一些研究认为活性氧(ROS)可能是在珊瑚白化中起到了重要作用(reviewed by Weis, 2008; Oakley & Davy, 2018)。按照这一氧化理论,高温及强光下共生藻的叶绿体光系统会接收到更多的能量,累积过量的电子,并且会生成更多氧气,高浓度的氧气则可以形成活性氧,共生藻中的活性氧可以向珊瑚细胞中扩散,珊瑚自身的线粒体也可在高温胁迫下形成活性氧。这样共生藻以及珊瑚中的活性氧可以在细胞层面造成各种氧化损伤,最终诱导珊瑚白化。然而新的单细胞层面上的研究却表明活性氧可能并不是珊瑚白化的初始引发因素(Nielsen et al., 2018),当高温胁迫已经引起一定程度白化时,珊瑚内的活性氧浓度并无显著上升,且珊瑚和共生藻中均未观察到明显的氧化损伤。
6 G0 r* Z- q9 o* F& R0 U( A
珊瑚可能失去共生藻的细胞机制包括藻类的原位降解、藻类被珊瑚宿主释放到胞外、含有藻类的宿主细胞脱离珊瑚体,以及含有藻类的宿主细胞凋亡或自噬(reviewed by Oakley & Davy, 2018)。按照Bieri et al.(2016)的研究,在高温或强光胁迫下,共生藻主要是通过胞吐或类似机制被珊瑚宿主完整地排到珊瑚细胞外。
- w0 k0 ^+ @- f6 Z 2.2 珊瑚对温度上升可能的适应
! f6 x6 }( ]( d/ T# `$ i' V& v
按照适应白化假说,不同的共生藻类群对温度(或其他环境因子)有不同的适应能力,不同温度下珊瑚可能可以调节其共生藻群落组成(e.g., Tong et al., 2017)。在环境胁迫下,珊瑚可以通过白化失去共生藻,随后从外界获得新的共生藻,以更好地适应环境。Baker(2001)进行的移植实验中,在从深水向浅水移植的珊瑚,在这种急性应激下虽然短期白化率较高,但随后其共生藻类发生了转变,珊瑚的长期存活率也较高;而从浅水向深水移植的珊瑚,虽然短期内没有经历白化,但其长期存活率较低,支持了珊瑚白化本身就是一种高风险的适应机制。野外的观察也表明,在高温导致的严重白化和死亡事件发生后,珊瑚中耐热的共生藻更为丰富(Baker et al., 2004; Jones et al., 2008)。Silverstein et al.(2015)的实验进一步确认了至少在某些情况下,珊瑚白化后的耐热性增加是由共生藻群落组成的变化导致的,而不是仅仅由于热暴露本身引起的珊瑚中的变化。
/ c# u; J) G" R7 P9 g3 l 除此之外,珊瑚还可通过共生藻转换以外的机制适应环境。生活在更热和更多变的泻湖中的风信子鹿角珊瑚(Acropora hyacinthus)对热应激白化的耐受能力更强(Oliver & Palumbi, 2011),并且将原本生活在温度较低处的风信子鹿角珊瑚移植到更热和更多变的条件后,珊瑚很快(1-2年)就获得了对热应激的耐受能力,这种环境适应应该主要是与珊瑚的基因表达变化有关(Palumbi et al., 2014)。更短期的热驯化(7-11天)也可提高珊瑚的耐热性,虽然此时在未接受热应激的驯化过的细枝鹿角珊瑚(Acropora nana)中未观察到基因表达变化,但驯化后的细枝鹿角珊瑚在热应激后表现出了广泛的基因表达变化,这表明珊瑚可能具有多级适应机制(Bay & Palumbi, 2015)。形态学层面也可观察到一些珊瑚的热适应变化,如热驯化可导致鹿角杯形珊瑚(Pocillopora damicornis)的胃层增厚,可能有利于保护共生藻免受过量辐射(Mayfield et al., 2013)。
0 f1 R9 K6 t0 Y! ^5 x4 ]4 h
但也有研究表明全球变暖可能会使得一些珊瑚的抗白化机制失效。Ainsworth et al.(2015)发现过去30年间过去大堡礁75%的热应激事件的温度变化情况都属于“保护轨迹”,即先有一个低于珊瑚白化阈值的亚高温,随后温度在平均约10天的回落期后再升高到白化阈值以上,这种情况下,前一个亚白化高温有助于珊瑚获得耐热性,从而降低珊瑚细胞的死亡以及共生藻的损失。但0.5 °C的全球变暖可能就足以使得热应激事件更多地从保护轨迹转变为单一白化轨迹和重复白化轨迹,即在达到白化阈值的高温事件前不再会有一个亚白化的高温,从而导致珊瑚的白化和死亡增加。
- A6 e6 ]- ?# \7 b& v0 X
2.3 全球珊瑚礁白化的时空格局
8 M/ S" X7 Y- a+ ?7 Q
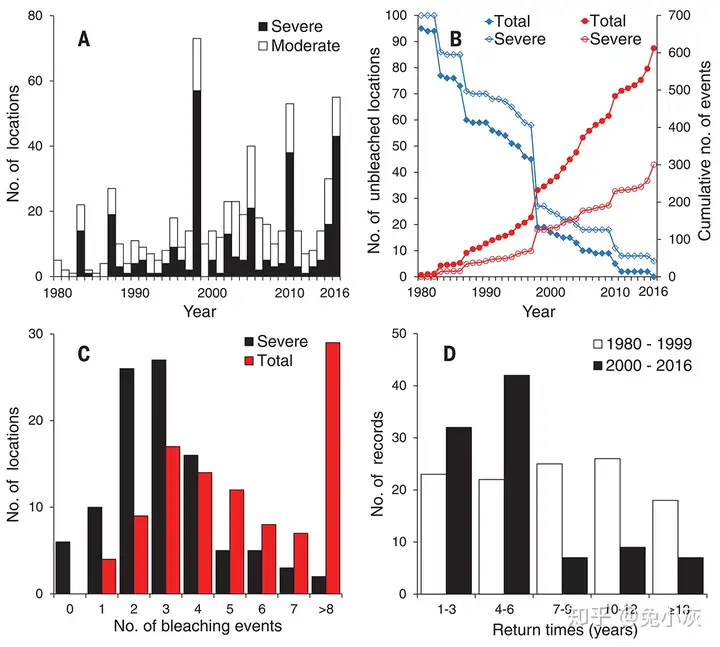
Lough et al.(2018)的统计表明从1871年至2017年全球范围内1670个热带珊瑚礁地点的平均温度已经上升了0.65 °C,略低于全球平均升温,但不同地点的变暖情况也有很大波动,一些地点的升温是高于全球平均情况的。热胁迫情况最严重的年份常与厄尔尼诺事件有关,2015-2016年的厄尔尼诺事件是有记录以来最严重的一次。自1980年以来,58%的严重白化事件发生在四个强烈的厄尔尼诺事件中(1982-1983、1997-1998、2009-2010和2015-2016),但随着全球整体变暖,可能在没有厄尔尼诺事件时夏季温度也可以容易地超过白化阈值(Hughes et al., 2018a)。
3 {: }/ o i( D1 K4 {
2015-2016年的白化事件影响了统计中75%的珊瑚礁,规模上与1997-1998年的事件相当。随着白化事件的地理覆盖范围不断扩大,珊瑚的潜在避难所也越来越少,在Hughes et al. (2018a)检查的100个地点中,只有6个尚未发生过严重白化。并且珊瑚礁在严重白化事件之间的恢复期也从1980年代早期的平均约27年急剧下降到2016年的约5.9年。
1 h/ V% X: m( y4 U( @* c1 K
随着全球变暖,强厄尔尼诺事件带来的热胁迫增加,导致全球性的珊瑚白化(引自Hughes et al., 2018a)在空间上,不同地区的白化历史也有所不同(Hughes et al., 2018a)。在20世纪80年代,西大西洋的白化风险最高,太平洋其次,印度洋和澳大利亚周边的白化风险最低。然而,在澳大利亚周边及中东地区,白化风险已经随着时间的推移迅速增加。
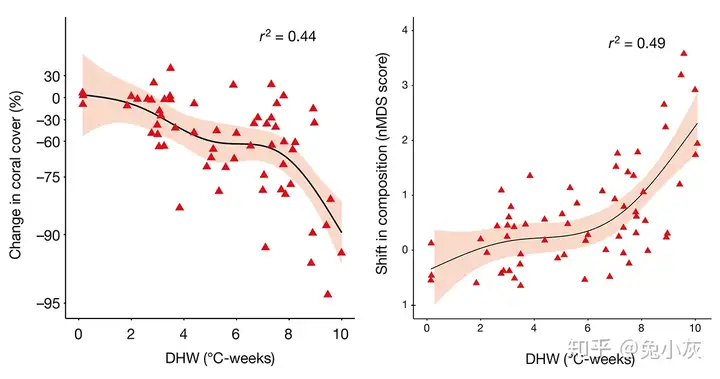
+ r' ]! L! t" z0 O 澳大利亚的大堡礁是一个目前关注的热点地区。从1985年到2012年,大堡礁硬珊瑚覆盖率从28.0%下降到了13.8%(Death et al., 2012)。1998年、2002年和2016年大堡礁经历了3次严重白化事件,在1998年中、南部的白化最为严重,2002年的白化区域更加广泛,2016年白化则还要更严重,并且北部受影响最强,珊瑚白化的这一空间模式可以很好地用热胁迫的空间模式来解释(Hughes et al., 2017b, 2018b)。对于热胁迫反复影响到的区域,珊瑚也经历了重复的白化事件(Hughes et al., 2017b)。另外热胁迫不仅可以解释短期的珊瑚白化现象,还可以解释更长期的珊瑚覆盖率下降(Hughes et al., 2018b)。在大堡礁的这3次白化事件中,没有发现前期白化对后续热胁迫下的珊瑚起到了保护作用(Hughes et al., 2017b)。在太平洋中部的Jarvis珊瑚礁在过去的60年间经历了8次严重白化事件,同样没有发现珊瑚适应的迹象(Barkley et al., 2018),这可能说明珊瑚的适应能力在野外实际情景下还需要进一步研究。
i2 n' ]* }. ] h: p& G
2.4 过度升温可能会抑制珊瑚钙化
+ B: W- {% n. X7 o! X# A5 l8 E. ]
尽管小幅变暖可能对珊瑚钙化有正面效应,但已经有研究认为安达曼海(Tanzil et al., 2009)和红海(Cantin et al., 2010; Steiner et al., 2018)中观察到的珊瑚钙化率下降可能与温度上升有关。事实上,珊瑚钙化中的pH调节过程依赖于共生藻的代谢活动,当温度上升至抑制共生藻乃至使珊瑚白化时,再考虑到海洋酸化的叠加作用,珊瑚的钙化过程也会受到严重抑制(McCulloch et al., 2017)。
% G1 N9 x3 n: t 2.5 温度上升会改变珊瑚礁系统的群落结构
' H6 ~* k; b( P 有多项研究表明关注了温度上升及其带来的白化事件会改变珊瑚的群落结构,某些珊瑚种类可能会严重受损,另一些种类的相对比例则增加(e.g., Loya et al., 2001; Harii et al., 2014)。Hughes et al.(2018b)对于大堡礁2015-2016年白化事件的研究表明较短期(8个月)珊瑚组合改变的程度与热胁迫呈正相关,严重白化(> 60%)的珊瑚礁中珊瑚组合发生了明显改变,其余珊瑚礁的改变则不明显。而日本的一项研究表明,长期上(10年后)虽然物种丰富度恢复到了白化前的水平,但珊瑚组合还是发生了变化,其主要成分包括了耐热的本地物种、能快速生长的本地残余物种,以及从附近重新迁移来的物种(van Woesik et al., 2011)。
! U$ ^/ p5 x$ g& ~0 w2 m0 a5 A 热胁迫引发的白化事件不仅可以导致珊瑚覆盖率下降,也会导致珊瑚群落结构发生变化(引自Hughes et al., 2018b)大堡礁2015-2016年白化事件后,在许多珊瑚发生衰退的珊瑚礁中藻类覆盖率发生了大幅增加,许多鱼类和无脊椎动物的空间分布格局也发生了变化(Stuart-Smith et al., 2018)。另一项研究则表明珊瑚的变化甚至可能会使某些鱼类的行为发生改变(Keith et al., 2018)。
" C% s: p5 m2 b9 J( g% r& [ 在长期的生态系统恢复过程中,塞舌尔的案例表明一部分白化后失去了> 90%的活珊瑚覆盖的珊瑚礁又恢复到了由珊瑚主导的状态,然而另一部分则没有恢复为珊瑚主导型,而是转变为了由肉质大型藻类主导的生态系统,Graham et al.(2015)认为这种演变轨迹的不同可能主要与水深以及珊瑚礁的结构复杂性等因素有关。
+ ?" R6 t3 ~7 k7 d2 o& J
在热胁迫带来的珊瑚白化过程中,珊瑚相关的微生物群落也会发生变化,温度上升时弧菌(Vibrio)或类似物种发生了增加,并且这种增加要早于肉眼可见的白化的出现;而在白化之后,细菌组合又恢复到了与白化前类似的情况(Bourne et al., 2008)。
2 t3 G; L6 K- q* s) {" Z 2.6 珊瑚礁的未来?
1 a( B) }: B3 B1 Y" a- J3 D 目前对珊瑚礁未来变化的预测还具有很多不确定性(Pandolfi, 2015)。有预测认为将全球变暖限制在2 °C能够有效延迟全球珊瑚严重受损的时间,并可能可以允许某些珊瑚有足够的时间适应环境(e.g., van Hooidonk et al., 2013)。然而,大多数这种预测都没有考虑区域性的气候以及局地的水动力条件,DeCarlo et al.(2017)认为对于许多浅水珊瑚礁生态系统而言,这些预测可能过于乐观。而且即便未考虑局地的水动力条件,也有相对不乐观的预测认为,如果要使全球多于15%的珊瑚礁受到保护,变暖就需要被限制在2 °C以内(Frieler et al., 2012)。
5 |, w) p# ^6 p o+ R& E" L
考虑到将全球变暖限制在2 °C(RCP2.6)可能已经很困难,Kwiatkowski et al.(2015)分析了向平流层注入气溶胶的地球工程手段对珊瑚礁的可能影响。平流层气溶胶注入在降温的同时也会使海洋酸化更为严重,不过按照Kwiatkowski et al.(2015)的估计,即便对于酸化敏感的珊瑚来说,这种地球工程对于减轻其白化可能也是有利的。
$ ` U) Q S# ~: r1 O2 D; l
3 海平面上升下的珊瑚礁
* w5 k# ~* K7 ^8 ?+ e) u& S/ H
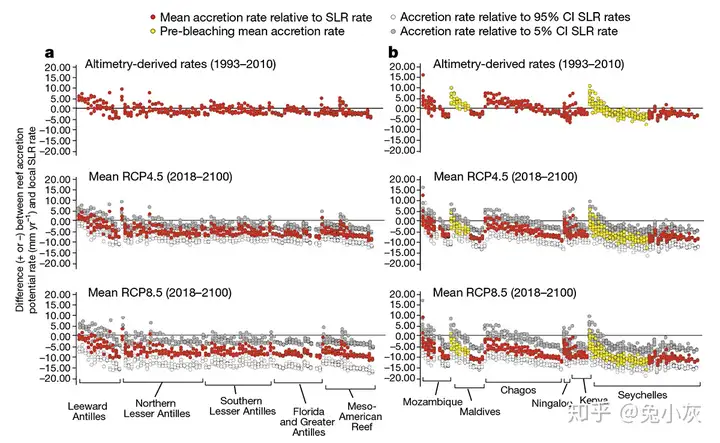
全球气候系统的变暖会导致上层海水热膨胀以及高海拔和极地地区的陆冰融化,从而引起海平面上升。关于珊瑚生长和海平面变化之间关系的研究还相对较少。有研究认为,至少某些珊瑚礁具有高的生长潜力,其增长能够与海平面的上升速率相匹配(Perry et al., 2015)。然而按照Perry et al.(2018)的研究,情况可能没有这么乐观,虽然在高珊瑚覆盖度下礁顶与海面的距离即使在RCP8.5的情形下也不会减小;但一旦珊瑚受到白化等事件的影响,覆盖度下降并且不能有效恢复,大部分珊瑚礁在RCP4.5下就无法跟上海平面的增长速度,而RCP8.5下到2100年礁顶与海面的距离则会增加0.5 m以上。无法跟上海平面上升的珊瑚最终可能会被“淹死”,珊瑚礁岛屿则可能更容易被侵蚀破坏(van Woesik et al., 2015; Woodroffe, 2008)。古海洋学的研究则表明在末次间冰期时发生过快速的跳跃式的海平面上升事件,数十年内海平面至少上升了2 m,墨西哥尤卡坦半岛的低位珊瑚礁在这次快速海平面上升事件中发生了珊瑚死亡和向陆后退(Blanchon et al., 2010)。
; o1 r) J' ]* l0 i o 在RCP4.5下,只有少数覆盖度比较高的珊瑚礁能够跟上海平面的增长速度(引自Perry et al., 2018)参考文献(就不列了吧……想搜应该都是搜得到的……)
0 t/ o1 _+ G6 o* e! @