7 e# d5 g2 ?) s/ f
# q) a' z' N1 N2 a , T) G5 M, i) m" R) }
3 c) u' ~3 f( i% T! f3 k" Z; I5 u- v( } 8 N1 c4 S8 p4 P) X7 p" D
沉降的海洋微粒中微生物硫酸盐还原和有机硫生成
! h; L4 O. w; i9 K; ?- [- B
/ _% x# a) s$ t, M1 G 气候的变化推动了海洋缺氧带(oxygen-deficient zone,缩写ODZ)的扩大,可能改变了全球碳、硫、氮和微量金属元素的循环。当前,对海洋缺氧是怎样影响有机碳的循环和埋藏还并不十分清楚,特别是在机理上。东热带太平洋和阿拉伯海的缺氧带约占海洋总量的0.35%,受人类活动造成气候变化的影响,它还在迅速扩大。缺氧带(ODZ)溶解的O2含量在 ~1 μmol/L以下,可能大部分低于10nmol/L。没有可利用的O2,动物基本被排除在外,微生物必须依靠替代的电子受体,如硝酸盐、三价铁和硫酸盐来呼吸有机质(organicmatter, 缩写OM)。一般来讲,只有电势更高的电子受体,如周围的硝酸盐消耗已尽才会发生生成硫化物的微生物硫酸盐还原 (microbial sulfate reduction,缩写MSR)。然而,地球化学和分子的证据都表明,尽管丰富硝酸盐存在,在缺氧带也会发生MSR。缺氧带活跃的MSR可能从根本上改变了其他元素的生物地球化学循环,因为生成的任何硫化物都可能迅速地与金属和有机质反应生成硫化物,这个过程称为硫化。
1 h$ K |2 \; v, y 由于硫化的潜在影响,近年来人们投入了大量精力来寻找缺氧带内活跃的微生物硫循环证据。在东太平洋、华盛顿边缘海和阿拉伯海的缺氧带检测到了硫循环的转录和基因序列,而在东北太平洋报导了镉丢失,因形成CdS。在圣胡安群岛大的海洋雪(marine snow)中检测到硫化物,通过放射性标记实验在智利离岸的缺氧带水体中测到低速率的硫化生产。然而,还没有人在微生物最可能代谢——大的沉降微粒检测MSR速率,探讨硫化物对沉积的OM组成有什么影响。Raven等人开展了这方面的研究,通过了解海洋缺氧带沉降中的微粒是否发生MSR和OM是否明显硫化,来探讨缺氧带扩大是怎样影响海洋中碳和金属的循环。
- _3 N. J' |) g4 ~0 ^ Raven等人在东热带北太平洋(the eastern tropical North Pacific,缩写ETNP)的缺氧带开展工作,包括浓缩沉降的微粒、加入放射性标记的硫酸盐一起培养和描述与任何生成硫化物有关反应产物的特征。2018年4月和5月间,他们通过R/VRoger Revelle考察船部署一个直径2米,50mm目的表面浮系微粒捕集器,4次采样:1次在P2(17.0°N×107.0°W,距墨西哥海岸200km,~3500m水深),没有大微粒控制,微粒通量相对低;3次在P1 (20.3°N×106.1°W,距岸~50km,~1500m水深),微粒通量相对高。收集的微粒是在深水次生亚硝酸盐浓度最高的位置(P1处为120到143m水深,P2处为147 m水深),那里硝酸盐还原会最强,因此也应是缺氧的OM循环速率最高的位置。将捕获的微粒在无氧条件下转移到血清瓶中,加入放射性标记的硫酸盐在黑暗中培养~24小时。然后,加入过量醋酸锌中止微生物活动。最后,分离出酸挥发的硫化物(AVS)(包括溶解的H2S和金属单硫化物),测定硫酸盐、AVS和耐水解固相等放射性。
8 ~0 u' z( M6 J8 v! \ + z9 G$ a* j7 B/ r
图1. 微粒培养中生成AVS和有机硫的放射性标记S积累。样品被沿x轴任意排列,y轴为对数。样品类型中AC为严格的缺氧收集器上采样,NW为净洗回收的。阴影组是单个部署的范例。至x轴的误差线表明结果与零没有显著差异。ND为没有检测到。 4 v/ y3 ^( k; y' D4 k
估计在培养过程中由MSR产生的大多数放射性标记硫化物会重新氧化,或者与金属或有机物反应生成固体物质。他们所测的AVS(图1)可能主要来自单硫化铁,而不是溶解的H2S。从处理方法看,在耐(酸)水解的残留固相中放射性标记的可能为元素S、有机S和/或黄铁矿,但他们后续的检测证明是有机S。P1的培养实验(平均,n=8)表明,24小时内累积171±10.8(2σ) nmol的AVS,或1711±100nmol的有机S;P2的实验只产生少量可见的团聚物,而且没有累积任何可分解的AVS(图1)。生成的有机S对AVS比例是变化的,这可能与当地沉降微粒中活性铁的含量有关。由此看来,传统的直接测定沉积物获得的有机硫含量也应该反映有机硫是MSR生成硫化物的主要成分。 4 l2 |2 u# ?' d6 `! i: v( E
可以根据这组实验推测原位S循环的速率,但需要知道那里微粒沉降的速率。在ETNP近岸微粒沉降速率大致为2m/day到120m/day,大部分情况都是慢的,沉降速率近6.5m/day。根据微粒捕集器的大小和孔目、微粒沉降速率和实测产物24小时的积累量可以计算MSR速率。如果沉降速率为6.5m/day,体积校正后P1处MSR的最小速率是~14pmol/L·hr。而高生产力且缺O2的智利边缘海的MSR速率(≤360pmol/L·hr)是P1处的~26倍,那里海洋表面的叶绿素-a浓度也比P1处高~22倍,它们的叶绿素-a浓度分别为~16mg/L和~0.7mg/L。因此,Raven等人得到的东热带北太平洋(ETNP)沉降微粒承载的MSR速率,按生产力比例换算后是可以与先前的Canfield的MSR速率大致进行对比。
- ?3 r% U5 I& A* @5 [: G 如果每个硫酸盐分子还原将使两个有机碳原子氧化,高通量的P1瓶中每天平均至少产生0.68μmol硫化物,这相当于每天有~1.4μmol有机C被呼吸掉。但瓶子中碳的确切质量不清楚。如果每瓶5mg C或另一测定结果~20 mg C,假定快速下沉微粒的有机碳转化速率为每天12%,则得到总呼吸每天消耗的有机碳分别为50μmol C和200μmol C,其MSR的呼吸掉的占总有机碳量的2.8%和0.7%。这与Bianchi等人预测ETNP缺氧带中有~1%有机碳被MSR呼吸消耗是一致的。
; a/ B9 x* a& e' i0 |! W
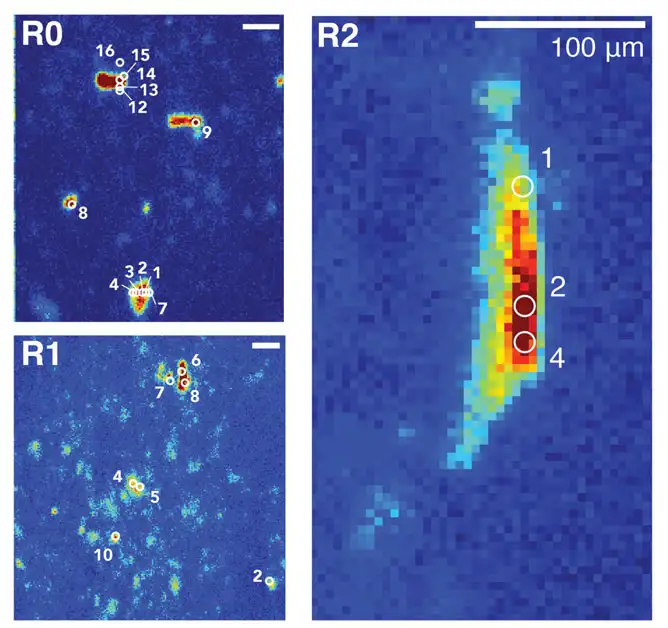
- U P3 t: [$ @ q 图2. 经过高温酸处理后超细玻璃纤维过滤器上沉淀的微粒。显示过滤器上所选扫描图区域0、1和2(R0、R1和R2)还原S的空间分布(2473.0 eV),颜色表示从低 (蓝色) 到高(红色)的相对吸收率。编号的白色圆圈显示采集完整光谱的位置。所有比例尺均为100μm,一个像素点呈现~5μm 2。
' R( ?7 @' |+ c5 ?9 ? u1 @/ S4 _) C 培养过程中沉降的聚集体为宏观的(>1mm),由与蓬松、近乎透明基质结合的一些暗色微粒构成。这些毛茸茸的基质加酸水解后溶解,原位留下两处离散的~100μm长微粒物,以及更多扩散的固体(图2)。可水解的基质中硫是相对氧化的,经K-edge X射线吸收光谱(XAS)鉴定是以硫酸酯和某些磺酸盐的形式出现(图3),这与硫酸多糖的存在一致。胞外聚合物中多糖丰富,硅藻和其他藻类会分泌出这类物质,海洋雪的形成与其有关。多糖外聚合物很可能是沉降微粒中硫化物池,因为硫化物可以将它们转化为更耐水解的形式。 + D8 [& z5 k i# {' T0 F9 v4 R
在离散、耐水解的微粒中的硫组分与水解的基质中所含大量的氧化态有机硫形成了对照。这些耐水解的S主要以有机硫化物形式存在,含少量的芳烃、二硫化物和更氧化态的。在过滤器内没有发现元素S、黄铁矿或硫酸盐的背景,这表明船上培养的放射性标记S产物主要是有机硫。扫描的R0和R2中离散的微粒主要与烷基硫化物一致(类型1为75.4±5.9%,平均值±1σ),而磺酸盐、酯类和二硫化物各自占百分之几。相比之下,R1中和R2中大颗粒内一个点(类型2)弥散物质更富集磺酸盐、硫酸盐酯和二硫化物,只含40.4%(±2.7%)的硫化物。
2 j% Q) g% U) g E 有机硫组分的空间差异可能反映了结合形成大的海洋微粒的OM来源和类型的异质性。有机硫化物来源包括在强酸水解和OM硫化反应都幸存的蛋白质中含硫的氨基酸。同样,磺酸盐和酯类可能基本表现海洋表层生命体的生物硫,像可水解的外聚物,或者它们也可以在OM硫化反应中与有机硫化物、二硫化物和芳烃一起产生。XAS的分析结果将之前的和现场培养的有机S整合到一起,因此任何这些有机S组合都可能是船上培养实验硫化产物的再现。但是,很清楚耐水解的OM可能是微粒承载的微生物硫酸盐还原期间生成硫化物的池。
1 j0 Q9 j; ~; O1 X/ W 在沉降微粒中硫化涉及到有机分子交叉接合和重排,这被认为是通过降低有机质与外酶的相容性来增强OM的保存。硫化程度体现在OM的S:C摩尔比值上,范围从新鲜的藻类生物的典型≤1%到许多黑色页岩的3%到5%。如果假设143m高通量收集器的每个瓶子中有原始5mg (20 mg) 的C (n=4) 在水解期间溶解,那么在一个~80到340μmol C的池中经过24小时的培养将有平均值为0.83μmol的有机硫积累。这样计算,不可能水解的OM的S:C摩尔比值每天增加~0.3%到1.1%,因为水体沉降几天之内就生成强硫化的OM(S:C≥3mol %)。因此,至少在足够还原和高微粒通量的情况下,沉降微粒好像是进入到海洋沉积物的一个可行硫化OM的来源。大多数可水解、未硫化的生物体随后在深埋之前,经过微生物作用再矿化,剩下硫化组分。因此,微粒承载的硫化能够为在海洋缺氧带之下所观测到的沉积物中高OM埋藏提供解释,而这缺氧带不是在沉积物-水界面之下。 % C! S# U. M& a4 {6 f- a1 G3 ?% M
在现在缺氧带(ODZ)的沉降微粒中所观测到的有机S组分也与已经发表大洋缺氧事件沉积的黑色页岩中古干酪根中的非常相似 (图3)。事件期间古北大西洋南部大部分地区都是硫化的,保存了特别富S的OM,这可能是沉降微粒快速硫化的结果。这些页岩中耐水解的有机S与现代缺氧带沉降微粒中的一样,主要由有机硫化物构成,其中二硫化物、芳烃和磺酸盐各自为~10%。两者的相似性可以解释为,现代缺氧带微粒中OM硫化的同样反应也在地球历史上更贫O2时期起到促进和提升OM的埋藏。因此,微粒承载的OM硫化是在缺氧条件下增强OM在沉积物保存的潜在机制。
- T) [8 f/ J( c4 c& _: H 6 \; r) B5 I4 f+ E6 W
图3. 东热带北太平洋(ETNP)沉降的微粒和白垩纪大洋缺氧事件2期间沉积的黑色页岩中的硫组分。用SIXPACK上线性去卷积拟合,色条宽表示不同硫组分的相对丰度。只显示高质量点的拟合 (X 2 <2)。酸化前的黑色页岩和微粒的色条图形代表所有可用数据的平均值。R0、R1和R2上测点位置如图2所示。 / L3 ]! w# q. ], Q& D* f& I8 }
现代缺氧带(ODZ)微粒承载的MSR和OM硫化的速率看来在时间上和空间上都是高度可变的,因此用它们来推算全球规模的有机碳埋藏还为时过早。然而,机理上看我们能够预测,海洋缺氧带(ODZ)的扩大会扩展沉降微粒为载体的OM硫化的规模。这将表现为气候和大气CO2的负反馈:提高CO2导致海洋缺氧带(ODZ)扩张,进而推动以OM形式沉积来增加CO2封存。采用现在的取样方法,开展更多研究,我们将可能量化全球海洋沉降微粒承载的MSR和OM硫化的规模,探讨它们对环境和气候的影响。
+ J" h; S% F( @ ) b; t: x9 A0 Z, M" O3 J) ^( U
参考文献1.J. Karstensen, L. Stramma, M. Visbeck, Prog. Oceanogr. 77, 331–350 (2008).2.N. P. Revsbech et al., Limnol. Oceanogr. Methods 7, 371–381 (2009).3.D. E. Canfield et al., Science 330, 1375–1378 (2010).4.M. R. Raven, R. G. Keil, S. M, Science 371, 178–181 (2021).5.E. L. Cavan, M. Trimmer, F. Shelley, R. Sanders, Nat. Commun. 8, 14847 (2017).6.D. Bianchi, T. S. Weber, R. Kiko, C. Deutsch, Nat. Geosci. 11, 263–268 (2018).7.M. R. Raven, D. A. Fike, M. L. Gomes, S. M. Webb, Front. Earth Sci. 7, 98(2019).8.M. R. Raven et al., Earth Planet. Sci. Lett. 512, 27–38 (2019).
$ e" P( v$ x& v2 ?6 ~, s7 Q4 Y (撰稿:储雪蕾)
+ K& C) q7 t# o0 ^% J+ v$ `5 G 校对:张崧 O5 R& v. U2 K
8 v. n7 S `% _1 l, Z
0 }$ T7 K; e$ U2 _9 p g7 ~7 y2 u5 a6 P C. o) t, \
|