$ X9 K/ m5 }: o0 F2 K* |, L$ ]5 R 7 F: e/ g& x/ J
# u9 r/ X5 W2 k0 M% A
{/ N2 O! N+ Z
2 K5 I. b: M, [- F. P0 U/ A 3 B- p' g% X! a, H1 R
1 I# N! B, v5 n; l( E |& t' n
! ]& L0 r0 d2 i/ r" u, y5 ^! o8 h 【前沿报道】Science:新生代海洋氮-硫循环变化及其与构造运动的关系
3 i/ A8 q4 T1 h* G
2 _8 Y! p) ~! j' P1 y; H3 M 海洋的氧化-还原状态决定着碳、氮、硫等元素的生物地球化学循环,与生命过程息息相关,是国际地学界长久以来的研究热点。目前,相关研究多集中在通过同位素手段来重建氮、硫循环过程的变化,并反演海洋的氧化和还原状态,进而探讨其与生命演替的关系。如埃迪卡拉纪(约6.35~5.42亿年)海洋氧化(Fike et al., 2006)与真核生物的起源演化、二叠纪末(约2.514亿年)海洋缺氧与生物灭绝(Schobben et al., 2015)等。然而,将氮、硫循环与构造运动相关联的研究尚属少见。实际上,构造运动会在更大尺度上决定着海洋格局变化,进而影响海水氧化还原和氮-硫循环过程,值得我们去关注。在这里以新生代早期构造运动对海洋氮、硫循环的作用为例,作以简要介绍。 " a7 n- R/ [$ W3 R$ T$ G1 s
2 ~$ o) L) H3 x+ x2 U
9 X% h8 m2 r& h' k- b/ F! F2 J6 { 3 D! w& M5 e5 {5 B6 k
海洋氮、硫循环过程主要表现为这两种元素的氧化态和还原态在水体中相对比例的变化。该比例变化取决于海水中氮和硫元素不同价态化合物的输入和输出情况。海洋中硝酸盐和硫酸盐的还原是影响两元素氧化态和还原态收支的关键过程,海水中氧气的多寡决定了海洋硝酸盐和硫酸盐被还原的程度。在次氧或贫氧环境的水体中,通常硝酸盐与硫酸盐还原菌较为活跃,致使反硝化(将NO3 -还原成N2)和硫酸盐还原(将SO4 2-还原成H2S)过程增强。由于细菌优先利用轻同位素( 14N和 32S),这两个过程均伴随着重同位素( 15N和 34S)富集在海水中剩余的硝酸盐和硫酸盐当中。但是,同位素变化最终是否能够被记录到还取决于还原反应中剩余的硝酸盐与硫酸盐的量。例如,在沉积物孔隙水中发生的反硝化过程,由于会将其中的硝酸盐消耗殆尽而不产生明显同位素分馏。 " U) p3 u( x6 F
3 Y7 F% |) Y. K2 B0 N
# A6 F# O8 \ |0 z9 A
) J2 D" L1 w) F! s+ O, y 目前,氮、硫同位素(δ 15N和δ 34S)分析是研究海洋氧化或缺氧状态的有效手段。在众多研究载体中,有孔虫壳体碳酸盐中的有机质被认为是分析氮同位素的可靠对象,而重晶石(BaSO4)和有孔虫碳酸盐晶格中的硫酸盐则是分析硫同位素的理想材料,主要是因为它们代表海洋来源且较少受到成岩作用的影响。本文讨论的研究成果均来自这两类物质的氮、硫同位素数据。
9 k9 M* Q6 w3 E- O: X* m8 S9 q
1 I' {5 o9 h9 h" i+ f2 F3 S ! o0 v# G8 c) [ D \% C
7 l6 b7 y( ?: j3 }: s
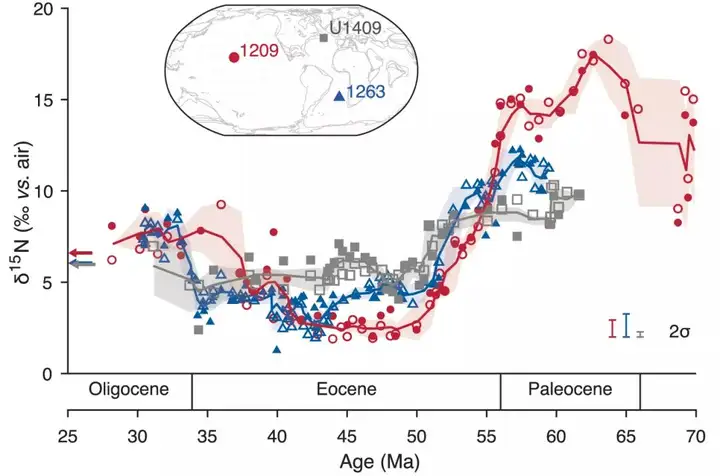
近期,美国普林斯顿大学地球科学系的Kast等在Science上首次报道了来自北太平洋(ODP 1209)、北大西洋(IODP U1409)和南大西洋(ODP 1263)新生代以来有孔虫壳体碳酸盐中有机质氮同位素(foraminifera-bound δ 15N, FB-δ 15N)记录,并讨论了与海洋O2含量相关的氮循环变化。在这里,FB-δ 15N被认为反映了海洋浅部亚表层(100-300m深)水体中硝酸盐的δ 15N变化。他们的结果显示:古新世时,FB-δ 15N值要比现在高出约5‰-10‰;古新世到早始新世(57-50 Ma)时,FB-δ 15N出现最大13‰的显著降低;随后,在中始新世以来,FB-δ 15N总体维持在在较低水平,尽管在渐新世时又有所增加(图1)。古新世时的FB-δ 15N值处于新生代来的最高水平,表明当时海洋硝酸盐的δ 15N高,可能反映了水体中反硝化过程与沉积物内部反硝化过程发生速率的相对变化,即具有较高的全球海洋水体反硝化速率,或者较低的沉积反硝化速率。 # k$ x, ~; R# i- @( L/ ?
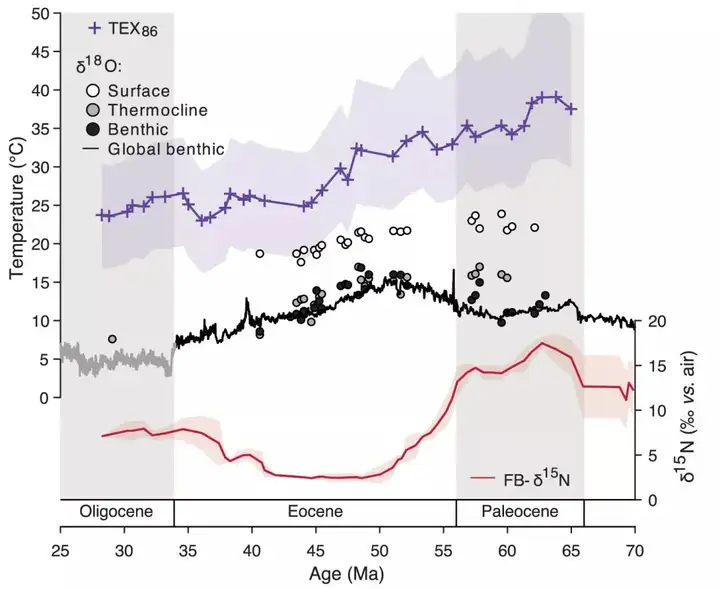
然而,FB-δ 15N表现出的空间差异(ODP 1209的δ 15N要高于ODP 1263和IODP U1409)更支持水体反硝化过程的增强,暗示古新世时海洋出现广泛的贫氧(suboxia)状态。此后,FB-δ 15N表现出显著降低,说明反硝化减弱和海水中氧气含量增加。由于始新世以来海水温度的下降(海水中氧的溶解度会随之增加)在步调上与此时δ 15N的降低不一致(图2),不能对其作以解释,作者将其归因于构造运动。在距今59 Ma时,印度和非洲板块向亚欧大陆的汇聚导致了特提斯洋的关闭,从此,温暖贫氧的特提斯洋水不再是太平洋中温跃层和中层水的来源,转而被寒冷富氧的高纬度表层海洋水所替代,从而导致水体贫氧状况缓解,反硝化强度减弱,δ 15N降低。
. }$ [3 L. u" V ! z q1 _3 ?7 c; Q' C8 v
图1 70 -30 Ma期间有孔虫壳体碳酸盐中有机质氮同位素变化。数据来自两种不同颗级有孔虫(颗粒>250 μm为实心,125-250 μm为空心)。实心线代表两种粒级数据的三点滑动平均,彩色背景区域为1σ不确定性区间。图中左侧箭头指示岩芯顶部的FB-δ 15N值:来自南大西洋516钻孔(蓝色)、U1409钻孔(灰色)和1209钻孔(红色)。插图为55 Ma前全球板块重建格局,表明本研究中ODP和IODP钻孔在当时的位置(Kast et al., 2019)
+ E* `; N0 k) k7 r* B. ~6 T
4 H/ r* [) W5 a1 B+ w' n 图2 北太平洋ODP 1209钻孔FB-δ 15N记录与温度替代性指标(TEX86和有孔虫δ 18O)的对比。底栖有孔虫δ 18O曲线的灰色段指示冰量变化可能对温度计算有影响(Kast et al., 2019) ' z# z6 ?* _7 Q0 c0 T& y
" p- I X: w5 [# a% J8 M; @( o
" z0 C1 g% B3 O# J. D* x
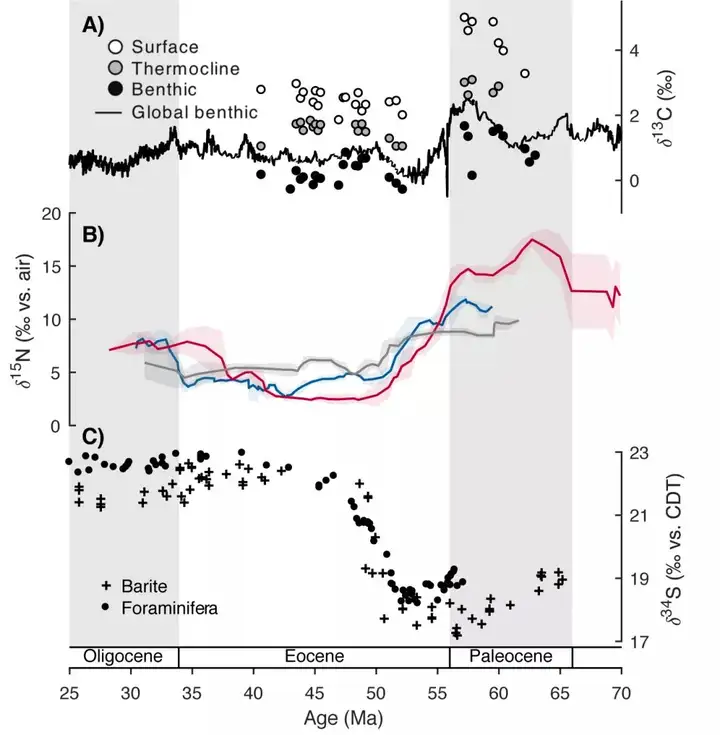
1 g4 F1 f i9 w# P 与此同时,海水中硫酸盐的δ 34S在古新世-始新世时段也经历了较大的变化。例如,Paytan等人早期在Science杂志上发表的重晶石δ 34S结果和Rennie等人在Nature Geoscience杂志上发表的有孔虫碳酸盐晶格硫酸盐δ 34S结果均记录了海洋硫同位素从53-40 Ma的显著升高,而且之后一直维持在相对高的水平(图3C)(Paytan et al., 1998; Rennie et al., 2018)。Rennie et al.(2018)认为构造运动所导致的硫化物埋藏海域的变化是上述硫酸盐δ 34S变化的主要原因。在新生代早期(约50.2±1.5 Ma),印度—洋内科希斯坦—拉达克岛弧发生碰撞,导致新特提斯洋的关闭和变窄,环特提斯洋边缘区域海平面快速下降,从而会导致浅海沉积环境的大规模减少。浅海沉积物中硫化物的δ 34S值通常要高于深海沉积物中的硫化物(因为浅层沉环境中硫酸盐微生物还原速率较快,同位素分馏较小),所以如果全球的硫化物更多地在深海环境中埋藏,势必会使硫化物δ 34S普遍降低,相应地海水中硫酸盐的δ 34S则会升高。
. K( ?; Z9 k) R6 v/ A) d5 H
3 G1 r5 m7 o, A* ^ 图3 新生代早期孔虫δ 13C值(A)与 FB-δ 15N记录(B)和重晶石、有孔虫碳酸盐晶格硫酸盐δ 34S记录(Paytan et al., 1998;Rennie et al., 2018)(C)的对比。从古新世到始新世,底栖有孔虫δ 13C值的总体下降可能表明有机质埋藏速率的降低,与大陆坡上方水体中较高的氧浓度是一致的(Kast et al., 2019, supplementary materials)
. J) m% U4 P) d1 d H& Z. {2 d4 @. Y
1 Y, `: I) \- ~, U% x, D$ {% M
2 e4 H: C2 N% ^6 v 这两篇文章均提出了新生代早期构造运动导致海洋环境格局的变化,从而分别改变了海洋氮、硫循环。但是,为了解释氮、硫同位素的各自特定变化,二者对构造变动时间和海洋格局变化的着眼点却不同,尚不能用一个统一的因素来解释氮、硫循环的变化。这体现了海洋中氮、硫地球化学循环影响因素的复杂性以及同位素研究的多解性,需要多指标综合分析研究。
+ S3 m8 T2 e* g5 W9 H 然而,Kast et al.(2019)提出的自59 Ma随着板块的汇聚海洋温跃层和中层水变得富氧,与Rennie et al.(2018)提出的构造变动导致硫化物埋藏至更深的海洋环境的观点并不是矛盾的。因为温跃层和中层水体的富氧会导致海洋硫酸盐还原界面向更深处移动,从而有利于更多的硫化物在深海环境中埋藏。值得注意的是海洋中溶解硫酸盐的滞留时间(~20 Ma)要远大于硝酸盐(<3千年),这至少是海洋硫酸盐δ 34S变化滞后于硝酸盐δ 15N约4 Ma的原因之一。今后的研究要综合多种替代性指标(如δ 15N、δ 34S和δ 53Cr等),同时注意各指标对驱动因素响应的时间尺度,采用模型分析获得更为确切的新生代海洋氧化-还原变化历史。
, p! Z' Y1 S6 J1 O, n : V' ?& ^+ w3 g* R! X6 L% d
主要参考文献 6 o- T h/ K& a/ q; Q" E/ U! c& j
1.Fike D A , Grotzinger J P , Pratt L M , et al. Oxidation of the Ediacaran Ocean[J]. Nature, 2006, 444(7120):744-747. " q- q2 L2 V+ c( ~) y, \; c. N
2.Kast E R, Stolper D A, Auderset A, et al. Nitrogen isotope evidence for expanded ocean suboxia in the early Cenozoic[J]. Science, 2019, 364(6438): 386-389. ; H2 R% ~% f/ L) q' W
3.Paytan A, Kastner M, Campbell D, et al. Sulfur isotopic composition of Cenozoic seawater sulfate [J]. Science, 1998, 282: 1459-1462.
9 i, i4 v) H; P 4.Rennie V C F, Paris G, Sessions A L, et al. Cenozoic record of δ34S in foraminiferal calcite implies an early Eocene shift to deep-ocean sulfide burial[J]. Nature Geoscience, 2018, 11(10): 761-765. ; y& Z- h+ z! u+ A- g% d" j
5.Schobben M, Stebbins A, Ghaderi A, et al. Flourishing ocean drives the end-Permian marine mass extinction[J]. Proceedings of the National Academy of Sciences, 2015, 112(33): 10298-10303. : N7 |8 M, c" n+ |. E
(撰稿:王旭/新生代室) % f$ g5 I$ `8 d$ x
美编:徐海潮 ( K' [2 |$ T" t
5 I5 Y$ p/ U7 v: ]
校对:HCN " B; o6 S9 t3 R' s
4 i o* `' [2 I; B2 R9 m/ D* R' @
( h4 z0 O9 [7 U: @" I
# ?0 C& T. p$ w, E) ? |