|
8 m l! V7 j% p% S
. H5 w; i0 n; |+ @ 发表期刊:The ISME Journal(IF=10.302)
+ ~7 r2 a3 d; V; J: l0 x3 I 发表时间:2020 ; K5 j$ Q k3 ]6 F
样本类型:水体
% R/ t3 F" x E; Y; d DOI:/10.1038/s41396-020-00825-6
; T. o$ d ], T 研究背景
7 @$ f0 ?! `# p) D( R5 R* x 病毒在海洋生态系统和生物地球化学中发挥着重要的作用。例如,裂解病毒可以针对微生物释放碳素,为微生物食物网提供食物来源(病毒分流),并对微生物群落组成产生直接影响。此外,编码辅助代谢基因(AMG)的病毒可以操纵它们的宿主,影响微生物的代谢和过程,如碳、氮、硫和铁循环。这些生物地球化学途径通常与环境条件密切相关,如水生生态系统中的氧跃层或化学跃层。
7 ]" e, t" N8 x3 m0 M: }3 A 实验设计 9 G' e* a* S# {7 D7 ^1 u
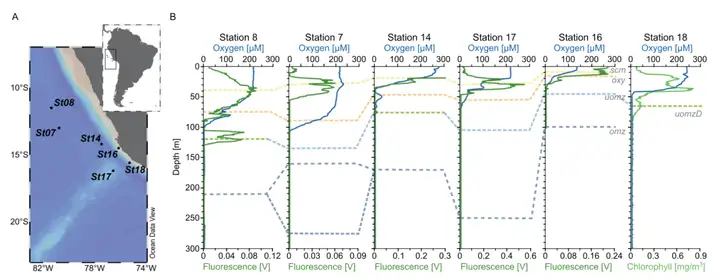
本研究在东热带南太平洋(ETSP)海洋氧最低区(OMZs)水域的六个深度剖面中,从跨越有氧到缺氧水域的22个样本中对总共29个病毒的宏基因组进行了测序(图1),研究结果评估了OMZ病毒是否可能通过AMG影响氮(N)循环。 / l8 D# |& q' n2 B4 f9 S' }* z) {
图1 研究区域概况和采样站的垂直特征。A 秘鲁海岸的ETSP最低氧区(OMZ)中7、8、14、16、17和18站点的位置。B 每个观测站的氧气(蓝色实线)和荧光/叶绿素(暗色和浅色的绿色实线)深度剖面主要结果 ( U" M% }3 o% i. e( _8 g3 D" z
1. ETSP样本宏病毒组数据分布及N相关AMGs
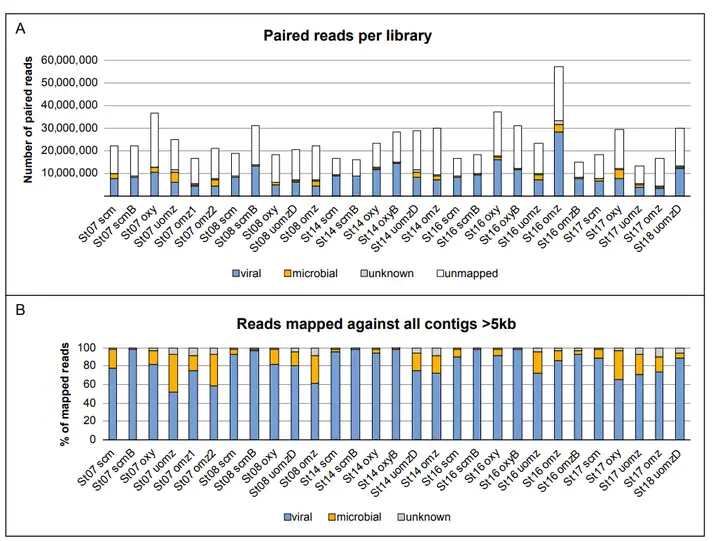
) w9 a* v" D# r+ b$ r$ t2 k: b 结果表明,29个宏基因组一共测序生成210 Gb数据,每个病毒组平均reads约49 M,并产生61,700个非冗余的长度大于5 kb scaffolds。其中,46,127个(75%)被鉴定为病毒,3,589个(6%)被鉴定为微生物,而其余11,984个scaffolds(19%)不能被明确鉴定(图2)。结果分析揭示了六种AMGs——focA、nirA、nirK、norB、amoC和glnK,其中由glnK编码的氮调节PII基因和由amoC编码的氨单氧酶是早前就被发现的两种AMGs。 5 b# D0 @0 W$ E8 r
图2 测序深度和获取的reads(1)铁氧化还原蛋白-亚硝酸还原酶和亚硝酸盐转运体 * y2 O4 ^5 x9 ]8 g* @& k
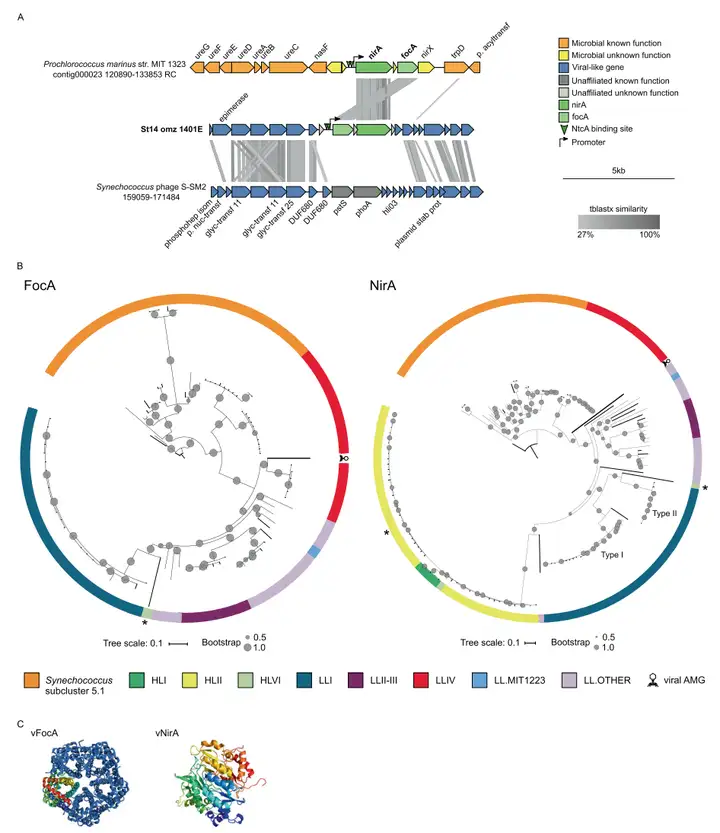
六个N循环AMGs中的第一个和第二个AMG包括一个亚硝酸盐转运蛋白基因(focA)和一个铁氧化还原蛋白-亚硝酸还原酶基因(nirA)。两个基因共定位在包含20个基因的病毒scaffold St14_omz_1401E(11,826bp)上,其中15个是病毒相关基因,两个预测NtcA结合位点,一个启动子,一个focA基因,一个未知基因和一个nirA基因,没有预测到终止子(图3A)。系统发育树分析表明,病毒FocA和NirA与原绿球菌和聚球藻属的物种最为相似,同源性分别为86%和67%(图3B),表明这两个AMG来源于这些宿主,并可能在感染过程中发挥有利作用,因为它们可以减少宿主争夺有限的硝酸盐和氨的需求。 , [2 {. Z( ~) T, p# s2 U
图3 病毒focA和nirA的基因组背景、多样性和蛋白质结构。A 编码nirA和focA的scaffold的遗传图谱及其与蓝藻和噬藻体参考基因组的比对。B 在ETSP和蓝藻序列中发现的病毒FocA或NirA的氨基酸比对得出的最大似然树。本研究中发现的病毒性AMGs的分支用粗线突出显示。C 病毒FocA的四级结构和病毒NirA的三级结构(2)亚硝酸盐还原酶和氧化氮还原酶 ) _$ Z' S" s/ c2 z W8 {! c
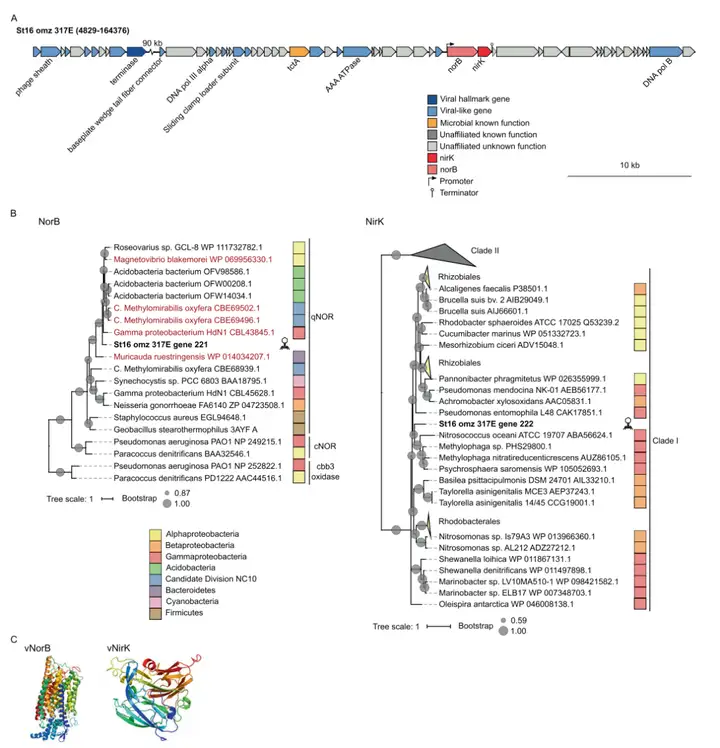
六个N循环AMGs中的第三个和第四个AMG包括一个含铜的亚硝酸还原酶基因nirK和一个氧化氮还原酶基因norB。病毒NirK蛋白与来自Clade I的微生物NirK聚集在一起,与Gamma变形菌纲最为相关(67-70%相似性),而NorB与酸杆菌门(57-62%相似性)、Magnetovibrio blakemorei(62%相似性)和Gamma变形菌纲HdN1(59.3% 相似性)最为相关(图4B)。在功能上,两种AMGs都是可能对病毒有益的反硝化基因,NirK蛋白将亚硝酸盐还原为一氧化氮,而NorB将一氧化氮还原为一氧化二氮,或一氧化氮还原为氮和氧。
* x/ l; w* }8 U) y* n 图4 病毒norB和nirK的基因组背景、多样性和蛋白质结构。A 编码norB和nirK的scaffold的遗传图谱。B 在ETSP和参考微生物序列中发现的病毒NorB或NirK的氨基酸比对得出的最大似然树。C 病毒NorB和病毒NirK的三级结构。(3)氨单加氧酶亚基C基因 X/ b1 t" t8 ^9 F3 Z
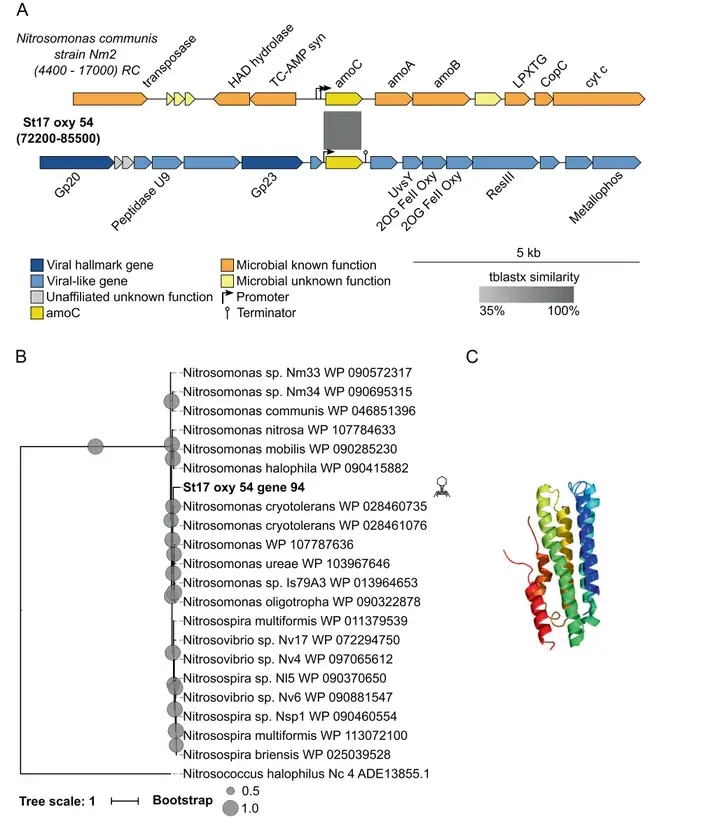
鉴定出的第五个AMG是氨单加氧酶亚基C基因(amoC)。以scaffold St17_oxy_54为代表的含有amoC的细菌病毒长度为144 kb,编码179个基因,两侧有一个启动子和一个终止子(图5A)。在进化上,amoC聚类在亚硝化单胞菌进化枝中(图5B),与来自亚硝化单胞菌的AmoC关系最密切(86%相似性)。在功能上,amoC可以通过获得氨氧化释放的能量来使OMZ病毒受益。
1 W) T; X/ _: p. F 图5 病毒amoC的基因组背景、多样性和蛋白质结构。A 编码类似细菌amoC的病毒scaffold的基因图谱,并与含有该基因的参考微生物基因组进行比对。B 从ETSP和参考微生物序列中发现的类似细菌病毒AmoC的氨基酸比对中得到的最大似然树。C 类似细菌病毒AmoC的三级结构。(4)GlnK-PII信号转导蛋白
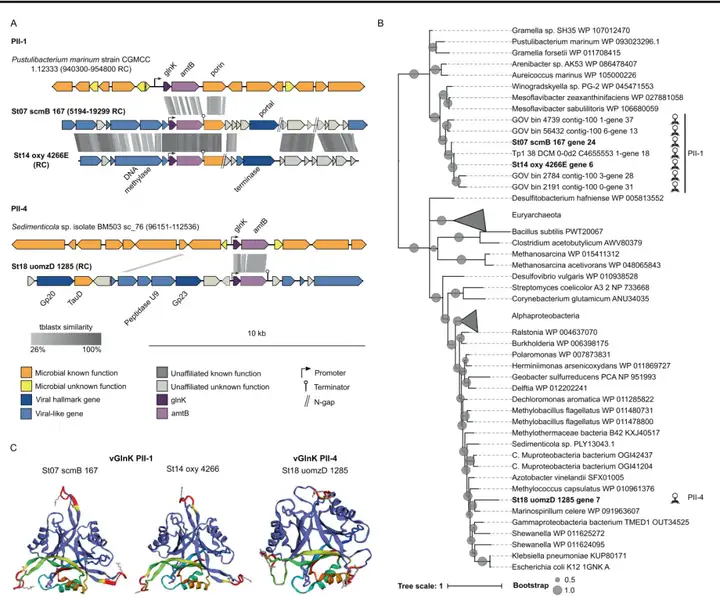
7 n3 A& I. b" k 六个与N相关的AMGs中的最后一个是GlnK,这是一种PII信号转导蛋白。本研究在29个宏基因组中共鉴定了三个含有glnK基因和amtB基因的病毒种群(图6A),前两个属于PII-1组(GOV数据工作中发现的glnK亚群的病毒代表),第三个是本研究指定为PII-4的新病毒glnK(图6B)。编码glnK的两个病毒scaffold上游都有一个启动子,铵转运蛋白基因(amtB)下游的一个终止子,以及紧邻amtB下游的孔蛋白基因 (ompL)(图6A)。在功能上,OMZ病毒可以通过调节宿主对氨的吸收而受益于glnK和amtB基因。 + n3 D, A5 L* {1 C# M
图6 病毒glnK的基因组背景、多样性和蛋白质结构 。A 编码glnK的scaffolds的遗传图谱,并与包含该基因的参考微生物基因组进行比对。B 在ETSP和参考微生物序列中发现的病毒GlnK氨基酸比对得出的最大似然树。C 病毒GlnK的三级结构。2. 病毒编码的N-循环AMGs的生物地球化学和生态学背景
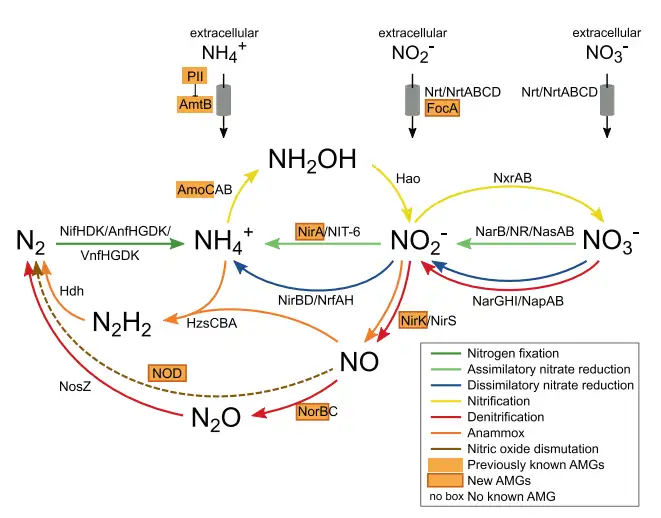
2 z* R4 ~% ~. D# K5 Y) s | 就N循环中AMGs来说,早先研究已经鉴定出硝化(AmoC)基因和氨调节(GlnK)基因,本次研究增加了编码介导同化亚硝酸盐还原(FocA和NirA)和反硝化(NirK、NorB或NOD-like NorB)基因。这些基因分散在已知的N循环途径中(图7)。 1 |# f* e& |0 n: P0 o
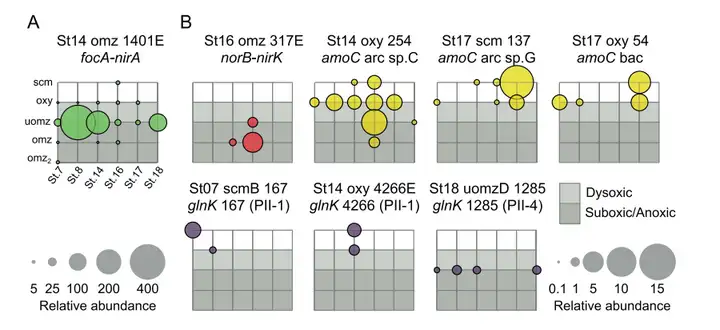
图7 病毒对氮气循环和运输的潜在贡献。该示意图代表了驱动氮循环以及参与的酶和转运蛋白的主要途径。含有病毒的蛋白质用橙色的背景颜色突出显示。与N循环相关的AMGs在ETSP OMZ不同站点的各个深度区的分布概况如图所示(图8)。含有focA-nirA的病毒种群,几乎存在于每个站点和每个深度,但在来自上层OMZ具有深叶绿素最大值的样本中要更加丰富。其他AMG编码病毒种群在整个数据集中的丰度较低,并且表现出强烈的深度偏好,例如,含有amoC的病毒种群主要分布在地表水和氧跃层水中,这在很大程度上遵循好氧氨氧化剂的分布(图8)。
7 ?9 f0 A0 x H 图8 含N-AMG的病毒种群在ETSP OMZ样本中的分布。气泡图代表了包含focA和nirA基因(A,绿色)、norB和nirK基因(B,红色)、amoC基因(B,黄色)和glnK基因(B,紫色)的病毒种群的相对丰度。研究结果表明,海洋氧最低区(OMZ)病毒不仅可以通过裂解关键的N循环微生物来影响N循环,而且还通过在感染期间调节不同的N代谢来影响N循环。随着这些N相关病毒AMGs被发现,未来的OMZ病毒工作可以评估病毒细胞影响的氮循环,差异量化病毒在OMZ N循环基因和转录中的生物地球化学影响。 5 Q7 X, n1 \ X& z% c* w& z3 ~
参考文献
" D4 x, W4 Z3 b Potential virus-mediated nitrogen cycling in oxygen-depleted oceanic waters. The ISME Journal, 2021. ( Q6 p/ \4 H5 j* q! a6 Y
DOI:10.1038/s41396-020-00825-6 $ @7 |) E, @ c) }9 F' t0 L' x
, ^; I' e7 O! M4 u
; b# v$ l b& b, ~' w, t
|