|
' o' c9 R r# m
Marine biofilms constitute a bank of hidden microbial diversity and functional potential
* q9 E7 r# n% o r+ E2 x 海洋生物膜隐藏的微生物多样性和功能性潜能
4 j+ K V$ a$ y3 u+ T 作者: Weipeng Zhang,Wei Ding, Yong-Xin Li等
+ z- G1 I7 a3 R% t- d ~9 ` 期刊: Nature Communications
. u" V4 m+ [% b+ |6 B 影响因子:11.878
" i% I# L. a# a4 m7 \3 Q 发表时间:2019-1 ; ^- i0 b4 O+ G) U
一、研究背景 / e) I% W( l. q; `
微生物在生物地理学循环过程、微生物之间的相互作用以及为脊椎动物提供重要的化学元素扮演着重要角色,维护并保持着海洋生态系统的健康和修复。随着高通量测序技术的蓬勃发展,海洋微生物的群落研究得到了空前的发展,比如著名的Tara Oceans 项目极大地促进了我们从全球角度去探索海洋微生物多样性。
3 u) E o- A2 A/ H9 j 但目前大部分的研究对象主要聚焦于海洋浮游微生物,全球大范围的海洋微生物微膜群落结构的研究还比较少。为了验证“海洋生物膜蕴藏着之前在海水中未有检测到的微生物,且它们有拥有着持续且迥然不同的重要功能”猜想,作者通过人造海洋生物膜结合高通量测序的方法对全球8个样点共101个生物膜样和24个海水样本进行测序,旨在全球范围里探索微生物膜“被隐藏”的微生物和功能潜能。
, M1 H8 ?) v* q4 M 二、实验设计
. e2 X+ m: O( n9 ?1 v9 T2 _ 本文作者在覆盖全球8个位置采样点里利用人工面板和聚苯乙烯培养皿在海水下1-2m制备生物膜,30d后采集金属表面形成的生物膜,共采集了101个生物膜样本以及临近样点海水样本24个,海水样本0.22μm和0.1μm滤膜器过滤后用DNA储存液储存跟生物膜样本一并于-80℃保存,生物膜样本同时用激光共聚焦显微镜进行结构观察。 , W' e+ G5 Q6 x! K- v+ ?
样本用试剂盒提取DNA后,建350bp小片段文库,用HiSeq 2500和HiSeq X Ten 进行宏基因组测序,生物膜和海水样本平均测序深度为115±39和130±33 milion reads。随后进行数据质量控制,用MEGAHIT进行reads拼接,同时下载了OM-RGC以及Tara Oceans项目中的宏基因组数据。
Y% d2 v. H8 P+ k) n; r! V 从宏基因组抽出16S 数据后预测出miTags,97%相似度聚类成OTU,随后对OTU进行了物种注释,为了得到测序深度对物种及鉴定的影响,把香港2017点11月采集的2个生物膜和海水样本进行深度测序(110Gb每样本),然后随机抽取特定数量reads进行分析。对所有样本进行了α和β多样性分析。 2 ?& z8 ~6 f) p0 @2 }. D; `
宏基因组数据随后进行了ORF预测,比对OM-RGC数据库,得到非冗余unigene集BCGC,用COG,KEGG,eggNOG,Pfam,SEED数据库进行功能注释。为了比较生物膜和free-living培养的微生物群落的区别,以某一香港海水样本来源的微生物进行室内培养实验,进行了群落结构分析。
( Z! v) _- g% A. i" D 最后,用Meta_RNA提取了16S rRNA gene序列,采用进化树的分析方法进一步挖掘新物种,同时抽取了长度大于10Kb的contigs用混合binning的方法做了基因组binning,检测了bins的完整度和污染度,用antiSMASH鉴定了生物合成基因簇基因,用MinCED挖掘了基因组的CRISPR信息。
; A# Z. l5 C3 S. b 三、实验结果 1、海洋生物膜的微生物群落和结构
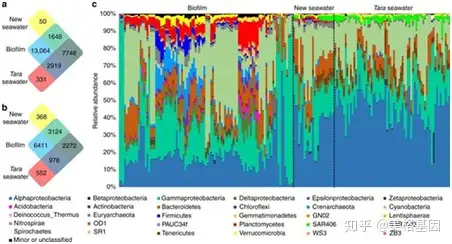
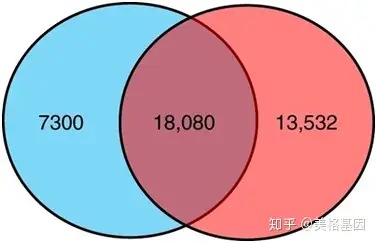
3 c5 L. V: X/ ]7 Y" e 16S群落结构分析发现,生物膜样本特有OTU有13,604,而新的海水样本有50个,Tara海水样本则是331个(Figure 1a),聚焦到丰富物种上(样本中每个OTU至少有9条序列),生物膜样本则有6411个OTU是特有的(Figure 1b);同时门水平的物种归属生物膜和海水样本有很大的区别(Figure 1c),和Tara的物种相比,生物膜样本中有7300个OTU在Tara样本中没有被检测到(Figure 2)。 4 y- I4 w, K. b% ]7 g
) k3 }+ j, D0 x; z' L Fig1 101个生物膜和24个海水样本微生物群落结构比较分析 7 J8 t i; e, e+ F
! i. h# C5 m9 w9 q3 r9 \ Fig2 生物膜样本和Tara样本OTU venn 图比较 5 T$ |9 f: x, F3 D7 A2 n5 a
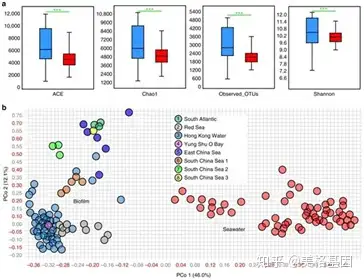
样本累计曲线分析发现,本次实验已经采集了足够多的样本和测序数据去解构微生物群落结构。抽取了10,000 miTags进行α和β多样性分析,生物膜和海水OTU中位数分别为2790和2155(Figure 3a),ACE、Shannon、Chao1显示生物膜的多样性高于海水样本,PCoA显示生物膜和海水样本的微生物群落结构能完全的分离开来。
8 J4 I5 J s! O* Z/ o: F6 `6 W- c % A- E( l* [; J! u$ X: _% \1 H
Fig 3 α和β多样性分析 " j- O0 I2 Y" U) x/ T9 j% Z, h
2、重要功能分析和实验依据
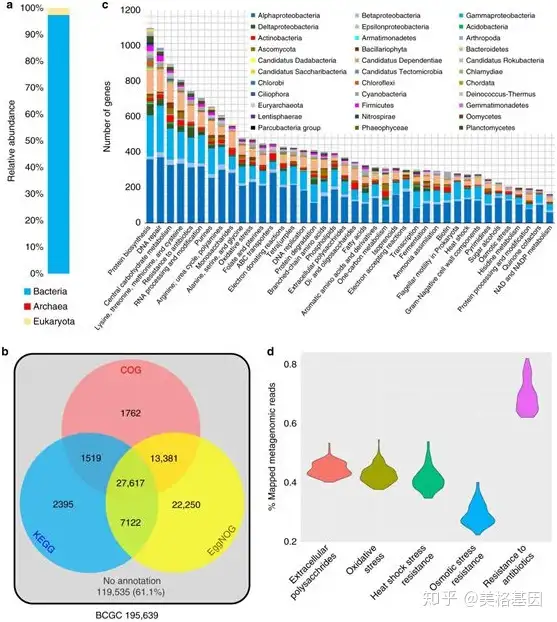
0 }5 B8 ~( }' g7 D) @( f) v( p 宏基因组数据分析发现生物膜样本里57.7%的ORFs在OM-RCS或新的海水样本gene catalog里没有比对信息,最后过滤找到了生物膜的core gene catalog(BCGC),共包含了195,639个非冗余编码蛋白基因。BCGC的物种注释发现97.3%的基因属于细菌,2.5%属于真核生物,古菌基因只有不到0.2%(Figure 4a),且只有38.9%的ORFs在COG,KEGG和SEED数据库里有比对信息(Figure 4b)。40个丰度最高的SEED功能类别包含了从生物膜形成、压力应答、环境适应到DNA修复等多个功能,这些基因横跨了众多的门中(Figure 4c)。Pfam的注释结果共找到了2567个结构域,这些结构域基因主要涉及压力抵抗和抗生素抗性,MEGARes比对结果从852个基因里共找到了18种抗生素抗性分类。 4 v. ~' s/ p3 ?( c
同时将香港的海水样本放于实验室进行free living和生物膜两种方式培养,该实验证了低丰度微生物从移动状态到表面附着的过渡期间,丰度低的物种及功能是富集的。
# V( L3 W; s; B, ?5 L7 u' [
: Y0 s6 ]% t" S6 \0 i3 ?. f2 } Fig4 biofilm core gene catalog(BCGC)的功能构成 % N X& a# H4 v" Z. Y X) c
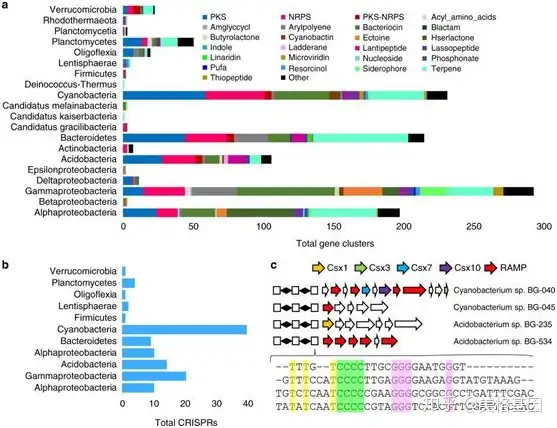
最后通过hybrid genome binning方法从宏基因组数据里组装到479个微生物基因组,它们分别属于25个细菌门,包括Alpha-、β-、γ-、δ-、ε-变形细菌等。基于binning的contig,共鉴定出了1148个生物合成基因簇,分类在了20个微生物门里(Figure 6a),其中分别有153、194和20个是合成NRPS、PKS和NRPS-PKS基因簇的。数量最大的基因家族来自γ-变形杆菌门,其次是蓝细菌门和拟杆菌门。同时鉴定基因组中的CRISPR arrays,共分布在了11个细菌门中(Figure 6b),最长的来自蓝细菌门,其次是γ-变形杆菌门和酸杆菌门。
9 F2 ?6 U! m7 K. a0 ?3 R3 O % p4 K& r1 h- F1 L
Fig5 479个微生物基因组(bins)的功能潜能分析
4 n; k' M1 H$ ?/ a4 L 四、结论 本文作者通过高通量测序对全球8个样点的101个生物膜样和24个海水样本进行高通量测序,结合Tara Oceans的宏基因组数据,研究了生物膜样的微生物群落结构。最终鉴定出了超过7300个在海水样本里没检测到的形成生物膜的物种,增加了对海洋微生物群落多样性的了解,为海洋生态位的分化提供了证据。基因功能鉴定发现了生物膜里重要的基因功能,包含了很多门的细菌,这些菌可能在压力应答、微生物互作上扮演着重要角色,Genome Binning也找到了很多新的生物合成基因簇和CRISPR-Cas系统。/ j8 s& q, \% v# A/ K2 b0 u
7 Q' d# j2 M$ l
4 Z& _' e0 q b3 k
- D! M) Q% C' s7 t l! K4 c& y. W
2 B! J4 k9 O# t
3 x" I4 G; u# _3 B% _& N
|